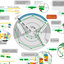
The evolution of oxygenic photosynthesis enabled organisms to use sunlight as an energy source, allowing them to colonize new niches. At the same time, life (as we know it) places severe constraints on photosynthesis. For example, the initial reactions of photosynthesis involve highly energetic intermediates that, if not controlled, can generate highly toxic side products (especially reactive oxygen species, ROS), that can damage other essential components of the organisms it powers. Photosynthesis must therefore be tightly regulated to balance the need for efficient energy conversion with the necessity of avoiding photodamage (Gust D, Kramer D, Moore A, Moore T, Vermaas W, Mater Res Bull 33:383–389, 2008). A related constraint on photosynthesis is the need to precisely balance how much energy is stored in ATP and NADPH to precisely meet biochemical demands. If this balancing does not occur, the system will fail, leading to photodamage (Kramer DM, Evans JR, Plant Physiol 155:70–78, 2011). Consideration of these requirements is essential for efforts to improve the efficiency of photosynthesis by introducing CO2 concentrating mechanisms, altering metabolism or biosynthetic pathways to shunt energy to alternative products (Kramer DM, Evans JR, Plant Physiol 155:70–78, 2011). These balancing processes must be extremely robust to contend with the rapid and unpredictable fluctuations in environmental conditions and metabolic demands that occur in nature. A large body of work has come from model systems, especially terrestrial higher plants and the green alga Chlamydomonas reinhardtii, leading to a model for the regulation of light reactions that involves 1) sensing of the pH gradient component of the thylakoid proton motive force (pmf), and 2) the redox state of the plastoquinone- and stromal pools. Over the short term, these sensors trigger regulation of light capture by altering the activity of ATP synthase leading to adjustments in lumen pH, which fine tunes light capture through nonphotochemical quenching (NPQ) and control of electron flow by adjusting the rate of PQH2 oxidation at the b6f complex. Simultaneously, this system controls the balance of ATP/NADPH by adjusting electron flux to linear and cyclic electron flow pathways to balance ATP/NADPH. This integrated “pmf paradigm” model explains much of the existing data on plants and green algae, but may not extend to other diverse organisms. This review considers how advances in our understanding of photosynthesis over the past 7–8 years, particularly in the discoveries of diverse biochemical/biophysical mechanisms in aquatic photosynthetic species, affects the view of energy balance, including the shunting of electrons to O2 through the flavodiiron proteins (FLV), the plastid terminal oxidase, the dissipation of electric field by ion movements, and the activation of alternative electron sinks. We will introduce the basic model that has been developed for higher plant chloroplasts, then contrast these with selected aquatic systems, focusing on how the differences impact the needs to re-balance both energy input and its partitioning into energy currencies.