Content uploaded by Weihong He
Author content
All content in this area was uploaded by Weihong He on Feb 28, 2016
Content may be subject to copyright.
This article was downloaded by: [British Geological Survey], [Mr James B. Riding]
On: 15 November 2013, At: 07:20
Publisher: Taylor & Francis
Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,
37-41 Mortimer Street, London W1T 3JH, UK
Palynology
Publication details, including instructions for authors and subscription information:
http://www.tandfonline.com/loi/tpal20
Latest Permian acritarchs from South China and the
Micrhystridium/Veryhachium complex revisited
Yong Lei a b , Thomas Servais b , Qinglai Feng a & Weihong He c
a State Key Laboratory of Geological Process and Mineral Resources , China University of
Geosciences , Wuhan , Hubei , China
b UMR 8217 Géosystèmes , CNRS-Université Lille 1 , Cité Scientifique, F-59655 Villeneuve
d’Ascq, France
c State Key Laboratory of Biogeology and Environmental Geology , China University of
Geosciences , Wuhan , Hubei , China
Accepted author version posted online: 15 Apr 2013.Published online: 04 Nov 2013.
To cite this article: Yong Lei , Thomas Servais , Qinglai Feng & Weihong He (2013) Latest Permian acritarchs from South China
and the Micrhystridium/Veryhachium complex revisited, Palynology, 37:2, 325-344, DOI: 10.1080/01916122.2013.793625
To link to this article: http://dx.doi.org/10.1080/01916122.2013.793625
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained
in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no
representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the
Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and
are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and
should be independently verified with primary sources of information. Taylor and Francis shall not be liable for
any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever
or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of
the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematic
reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any
form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://
www.tandfonline.com/page/terms-and-conditions
Latest Permian acritarchs from South China and the Micrhystridium/Veryhachium
complex revisited
Yong Lei
a,b
*, Thomas Servais
b
, Qinglai Feng
a
and Weihong He
c
a
State Key Laboratory of Geological Process and Mineral Resources, China University of Geosciences, Wuhan, Hubei, China;
b
UMR 8217 G
eosyst
emes, CNRS-Universit
e Lille 1, Cit
e Scientifique, F-59655 Villeneuve d’Ascq, France;
c
State Key Laboratory of
Biogeology and Environmental Geology, China University of Geosciences, Wuhan, Hubei, China
Diverse and well-preserved latest Permian phytoplankton assemblages are described from four sections of the
Yangtze Block, South China from localities in Zhongzhai (Guizhou Province), Shangsi (Sichuan Province),
Xiakou (Hubei Province) and Dongpan (Guangxi Province). Most of the species have been reported previously from
other upper Permian sections worldwide. The South Chinese phytoplankton taxa are generally very small in size,
usually displaying diameters of about 20 mm and commonly include the genera Micrhystridium,Veryhachium and
Leiosphaeridia. However, larger taxa with vesicles often exceeding 80 mm in diameter, such as Dictyotidium, are also
abundant in the Shangsi section. Due to the presence of large populations of Micrhystridium and Veryhachium,a
simple classification scheme for the Micrhystridium/Veryhachium complex is proposed, based on the geometrical
shape of the vesicle. We propose dividing the complex into five groups: the Veryhachium cylindricum group,
representing all ellipsoidal specimens; the Veryhachium trispinosum group, all with triangular-shaped vesicles; the
Veryhachium lairdii group, all with rectangular forms; the Micrhystridium pentagonale group, all with pentagonal
specimens; and the Micrhystridium breve group, which includes all spherical forms.
Keywords: phytoplankton; Micrhystridium/Veryhachium complex; Late Permian; Yangtze Block; China
1. Introduction
The group Acritarcha was created by Evitt (1963) as an
informal grouping of all organic-walled microfossils with
unknown or uncertain biological affinities (Servais 1996;
Strother 1996). They first appeared in the Proterozoic
and reached maximum diversity during the early and
middle Palaeozoic, especially during the Ordovician and
Early Devonian (Strother 1996; Servais et al. 2004,
2008). However, acritarch diversity dropped dramati-
cally in the late Palaeozoic, particularly between the Late
Devonian and Carboniferous, with very low diversities
in the Carboniferous and Permian, leading to almost a
complete absence of organic-walled microplankton, that
was named by Riegel (1996,2008) the ‘Phytoplankton
Blackout’. Servais et al. (2006) questioned the existence
of a ‘phytoplankton blackout’, because the absence of
resting cysts in the fossil record does not necessarily
imply the absence of phytoplanktonic cyst-producing
organisms in the late Palaeozoic oceans. Subsequently,
Mullins and Servais (2008) reviewed the diversity of the
Carboniferous phytoplankton, and noted that phyto-
plankton diversity was still high in the earliest Carbonif-
erous, but declined significantly from the Tournaisian to
the Serpukhovian. Phytoplankton diversity in the Late
Carboniferous was generally very low, with typically
only one to three species being documented in each
assemblage (Mullins & Servais 2008). Subsequently, Lei
et al. (2013) similarly reviewed the diversity of the phyto-
plankton of the Permian, and pointed out that phyto-
plankton diversities were much more higher than
previously assumed, with highest diversities in the Early
Permian and the latest Permian, where more than 30
phytoplankton genera have been reported.
Chinese acritarch studies, like elsewhere, have
focused mainly on the early and middle Palaeozoic, with
only a few studies in the Permian (Li et al. 2002,2004).
In the Late Permian of South China, most palynological
investigations were focused on the taxonomy and
biostratigraphy of spores and pollen grains. Only four
papers (Ouyang 1982,1986; Ouyang & Utting 1990;Li
et al. 2004) have described acritarchs from the Permian–
Triassic boundary (PTB), documenting the presence of
13 genera of acritarchs and related forms: Archaeodi-
nium,Baltisphaeridium,Cymatiosphaera,Dictyotidium,
Leiosphaeridia,Micrhystridium,Psiloschizosporis,Very-
hachium,Reduviasporonites,Schizosporis,Solisphaeri-
dium,Tasmanites and Tunisphaeridium. In these studies,
the small taxa (with diameters ca. of 20 mm, such as
Micrhystridium,Veryhachium and some species of Leios-
phaeridia) are the most common.
*Corresponding author. Email: leiyongdida@163.com
Ó2013 AASP – The Palynological Society
Palynology, 2013
Vol. 37, No. 2, 325–344, http://dx.doi.org/10.1080/01916122.2013.793625
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
This paper documents the Late Permian acritarch
populations from four sections of the Yangtze Block,
South China, with special focus on the dominant group
of taxa belonging to the Micrhystridium/Veryhachium
complex, for which we propose a simple classification
scheme of five informal groups.
2. Geological setting
The Yangtze Block of South China displays numerous
continuous sections of Upper Permian and Lower Tri-
assic strata from different palaeoenvironments, rang-
ing from nearshore to platform and slope settings
(Figure 1C). In the present study, four sections have
been investigated in the Guizhou, Sichuan, Hubei and
Guangxi provinces (Figure 1A, B). For each of these
sections, the reader is referred to the relevant geologi-
cal and palaeontological studies.
2.1. Zhongzhai section
The Zhongzhai section (2609.110N; 10517.113E) is
located about 5 km north of Zhongzhai Village, near
Langdai Town, Liuzhi County, Guizhou Province
(He et al. 2008). The section is located in the western
Yangtze Block (Figure 1); the uppermost Longtan
Formation (beds 1 to 27) and the lowermost Yelang
Formation (beds 28 to 37) are investigated in this
study. The Longtan Formation is mainly composed of
brownish sandstones and mudstones with a few lime-
stones. The lowermost Yelang Formation is dominated
by yellowish green or purplish red silty calcareous
mudstones, and a few limestones and claystones at its
base. According to the conodont zonation, Metcalfe
and Nicoll (2007) considered that the PTB in the
Zhongzhai section is in bed 28 (Figure 2). Based on
uranium-lead (U-Pb) dates, the absolute age of the
claystone in bed 29 indicates an age of 252.24 0.13
Ma (Shen et al. 2011).
2.2. Shangsi section
The Shangsi section is located near Guangyuan City in
the northern part of Sichuan Province, and belongs to
the northwestern corner of the Yangtze Block (Lai
et al. 1996; Figure 1). Both the Dalong (upper Wuchia-
pingian to Changhsingian) and the Feixianguan forma-
tions (Induan) are exposed here. The geology of the
Shangsi section was first described by Li et al. (1986).
Beds 17 to 22 are dominated by interbedded carbona-
ceous-siliceous rocks and siliceous limestones with a
few mudstones. The interval from beds 23 to 29 is
Figure 1. Location of the four sections studied in South China (Zhongzhai section, Shangsi section, Xiakou section and
Dongpan section) (A and B), and the latest Permian palaeogeography of South China (C, modified after Feng & Gu 2002).
326 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
mainly composed of siliceous mudstones and calcare-
ous mudstones with few claystones. Recently, Jiang
et al. (2011) suggested that the PTB be placed 22 cm
above the base of bed 28, based on hindeodid and
gondolellid conodont taxa. Similarly to the Zhongzhai
section, an absolute age (252.28 0.13 Ma) was deter-
mined from the claystones of bed 27 by Shen et al.
(2011).
2.3. Xiakou section
The Xiakou section (3106.874 N; 11148.221E) is situ-
ated near the town of Xiakou, in Xingshan County,
Yichang City, Hubei Province. It is located in the north
of the Yangtze Block (Wang & Xia 2004; Figure 1) and
comprises the uppermost Changxing Formation (beds
223 to 256), which is dominated by mudstones and
muddy limestones, and the lowermost Daye Formation
(beds 257 to 270), which is dominated by mudstones
with rare claystones. According to the conodont biozo-
nation, the PTB is placed in bed 262 (Shen et al.
2012a), rather than in bed 266 (Wang & Xia 2004;
Hong et al. 2008,2011).
2.4. Dongpan section
The Dongpan section, located at 2216.196 N and
10741.505 E (Shen et al. 2012b), is situated 80 km
southwest of Nanning, Guangxi Province, in the south-
ern area of the Yangtze Block (He et al. 2007b)
(Figure 1). Here, the Dalong Formation (beds 2 to 12)
and the Luolou Formation (beds 13 to 14) are exposed.
Figure 2. Phytoplankton distribution in the four sections investigated.
Palynology 327
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
The interval from beds 2 to 8 is mainly composed of
well-bedded siliceous rocks, muddy siliceous rocks and
mudstones. The interval from beds 9 to 14 is
dominated by mudstones and silty mudstones with
thin claystone intercalations (Feng et al. 2007).
According to He et al. (2005,2007a,2007b) and Feng
et al. (2007), beds 2 to 12 are late Changhsingian, and
bed 13 is considered to be early Induan.
3. Materials and methods
Two hundred and sixty nine samples were collected
from the four sections (45 samples from the Zhongzhai
section, 141 samples from Shangsi, 60 samples from
Xiakou and 23 samples from Dongpan). All of the
samples were processed for palynological analysis.
Fifty grams of each sample were prepared using
standard palynological methods. After treatment with
hydrochloric acid (33%) and hydrofluoric acid (40%),
the samples were concentrated by treatment with zinc
bromide solution (s.g. 2.2). The samples were not
oxidised or sieved. The residue was studied using light
microscopy and scanning electron microscopy (SEM).
All residues are stored in the collections of the China
University of Geosciences, Wuhan.
4. Results
The present investigation indicates that diverse and
abundant phytoplankton occur in the latest Permian of
the Yangtze Block, South China, from the four sec-
tions investigated (Figure 2). The highest diversity is
observed in the Shangsi section, with 20 species and
eight genera recorded in 37 samples, including Balti-
sphaeridium sp., Dictyotidium reticulatum,D. sp., Leio-
fusa sp., Leiosphaeridia microgranifera,L. minutissima,
Micrhystridium breve,M. fragile,M. pentagonale,
M. stellatum,Reduviasporonites catenulatus,R. chalas-
tus,Schizosporis scissus,S. sp., Veryhachium cylindri-
cum,V. hyalodermum,V. nasicum,V. quadratum,
V. sedecimspinosum and V. sp.
In the Zhongzhai and Xiakou sections, only a few
species occur in several samples, including Baltisphaeri-
dium sp., Dictyotidium reticulatum,Leiosphaeridia
microgranifera,Micrhystridium pentagonale,M. stella-
tum,Reduviasporonites catenulatus and R. chalastus.
In addition, only two species of Leiosphaeridia occur in
the Dongpan section. However, they are abundant
in 15 samples (Figure 2). The palynofacies changes in
the Shangsi section are described by a Lei et al.
(unpublished data), and the palaeoenvironmental dis-
tribution of the different was phytoplankton taxa
interpreted by (2012). The present paper describes
the systematic palaeontology of the assemblages
investigated, with special focus on the Micrhystridium/
Veryhachium complex.
5. The Micrhystridium/Veryhachium complex revisited
The genera Micrhystridium Deflandre 1937 and Very-
hachium Deunff 1954 are amongst the most widely
recorded acritarch genera throughout the entire
Phanerozoic. Fensome et al. (1990) listed more than
200 species of them. With a wide range of diagnoses,
they became typical ‘waste-basket’ genera, similar to
other genera such as Baltisphaeridium (Eisenack 1958)
Eisenack 1969,Multiplicisphaeridium Staplin 1961, and
Polygonium Vavrdov
a1966 (see Servais et al. 2007).
Many palynologists described new species of small
spherical acanthomorth acritarchs and placed them in
Micrhystridium, whereas numerous new small polygo-
nal acritarch species were attributed to Veryhachium.
However, because the interspecific variability is very
great, it is commonly difficult if not impossible to dis-
tinguish the different species, as well as differentiating
the two genera. These genera and their species thus
form a large plexus with a continuum of intermediate
morphotypes.
Generally, the Permian specimens attributed to
Micrhystridium display a spherical or oval vesicle,
bearing many proximally open spines with closed tips,
most often simple, and rarely branching. The vesicle
diameter is usually less than 20 mm (larger species were
usually attributed to the genera Baltisphaeridium
and/or Multiplicisphaeridium). However, Veryhachium
embraces all smaller acritarchs with polygonal body
outlines and less than eight spines.
In the Permian, the two genera Micrhystridium and
Veryhachium are very common. According to a recent
literature search, 27 species of Micrhystridium
were recorded 55 times in 26 publications, whereas
22 species of Veryhachium were documented 46 times
(Lei et al. 2013). At the species level, Micrhystridium
breve,M. stellatum and Veryhachium reductum are the
most frequently reported species of the Micrhystri-
dium/Veryhachium complex (Lei et al. 2013).
The genera Micrhystridium and Veryhachium have
a complex taxonomic history and both have been
revised and reviewed by many authors (see Sarjeant &
Stancliffe 1994; Servais et al. 2007). The taxonomy is
still being debated, and it remains difficult to identify
some of the individual species. This problem of identifi-
cation was noted very early, and some authors used the
term ‘Micrhystridium/Veryhachium complex’ several
decades ago (e.g. Wall & Downie 1963; Visscher and
Brugman 1981; Sarjeant & Stancliffe 1994; Eshet et al.
1995; Li et al. 2004; Van de Schootbrugge et al. 2005;
Riegel 2008).
328 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
It is not the objective of this paper to fully review
the classification scheme of the two genera, or to
emend their diagnoses which have been modified sev-
eral times already. A full revision, including population
analyses with biometric studies, of all described taxa of
the genera Micrhystridium and Veryhachium is neces-
sary in order to understand the taxonomy of these
taxa, and to provide synonymy lists. Our objective is to
propose a simple classification scheme that can easily
be used for analyses of latest Permian phytoplankton
assemblages, especially by palynologists who do not
describe the acritarch taxonomy in detail. However,
the simplified classification scheme presented here does
not consider all previously described taxa as synonyms.
In the present study of Late Permian phytoplank-
ton assemblages from South China, 10 morphotypes
within the Micrhystridium/Veryhachium complex are
recognised, and are here tentatively attributed to for-
merly described species. However, interspecific and
intergeneric variability is large and it is difficult to
draw clear lines between species. For identifying this
complex more easily, we propose to divide them into
five informal categories, which are based on the geo-
metric shape of the vesicle and do not take into account
the morphologies of the spines (or appendices)
(Figure 3).
Servais et al. (2007) revised the oldest morphotypes
of Veryhachium that first appeared in the Ordovician
and, following the informal usage of several authors,
proposed two informal groups for the genus Very-
hachium in order to facilitate their classification. The
Veryhachium trispinosum group was proposed for tri-
angular specimens, and the Veryhachium lairdii group
for rectangular forms.
Compared to the Ordovician, the Permian morpho-
types of Veryhachium are even more variable and
include morphological transients that range into
Micrhystridium. Therefore, we continue the reasoning
of Servais et al. (2007) and propose three additional
informal groupings to facilitate the classification of
these morphotypes in the late Palaeozoic. The informal
Veryhachium cylindricum group is here proposed to
include all ellipsoidal specimens of Veryhachium,
whereas two informal groups for Micrhystridium are
Figure 3. Idealised line drawings of the simplified classification scheme of the Micrhystridium/Veryhachium complex.
Palynology 329
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
proposed. These correspond to the Micrhystridium pen-
tagonale group for all pentagonal and hexagonal speci-
mens, and the Micrhystridium breve group for all
spherical specimens.
In our study, the Veryhachium cylindricum group
includes the previously described species Veryhachium
cylindricum (Table 1). The Veryhachium trispinosum
group includes all triangular specimens, amongst them
Veryhachium hyalodermum and Veryhachium sp. The
Veryhachium lairdii group includes all morphotypes
with rectangular or square vesicles, such as
Veryhachium nasicum,V. quadratum and V. sedecimspi-
nosum.TheMicrhystridium pentagonale group includes
the two species Micrhystridium pentagonale and
M. stellatum, whereas the Micrhystridium breve group
includes both Micrhystridium fragile and M. breve.Itis
clear that this proposal is a tentative classification into
an informal subgeneric scheme, and other geometric
shape-like groupings could be added if necessary. The
idealised line drawings of Figure 3illustrate that all vesi-
cle shapes from triangular to square, pentagonal to
polygonal, round and ellipsoidal may exist, and Plate 2
shows that morphotypes with such vesicles shapes are
actually present in the Permian assemblages. It is obvi-
ous that these specimens belong to a Micrhystridium/
Veryhachium complex, and that the determination at
the specific level becomes problematical when dealing
with larger populations, such as those recorded from
the Chinese Permian.
The objective of this tentative classification into
informal groupings is simply to propose a rapid classi-
fication scheme of the Micrhystridium/Veryhachium
complex. We do not know if these morphotypes actu-
ally represent the cysts of a single biological species, or
of a greater number of taxa. It is beyond the scope of
this paper to answer that question. However future
research, such as analysis of the biomarkers or
biochemistry of the vesicle walls, would help facilitate
a better understanding of the generic relationships of
the different taxa. Interestingly, Grice et al. (2005) and
Hays et al. (2011) analysed the biomarkers of sedi-
ments from Greenland at the Permian–Triassic bound-
ary, and suggested that the C33 n-alkylcyclohexane
may be a potential biomarker for the Micrhystridium/
Veryhachium complex. So far, these data result from
the analyses of both Micrhystridium and Veryhachium.
Future research of separated Micrhystridium and Very-
hachium populations may provide more precise
interpretations.
6. Systematic palaeontology
Acritarchs are, by definition organic-walled microfos-
sils of unknown biological affinities (Evitt 1963; Servais
1996). Several authors have suggested, mostly based on
morphological comparisons with extant phytoplank-
ton, biological relationships with various microalgal
groups. According to various authors (e.g. Colbath &
Grenfell 1995;LeH
eriss
e et al. 2009), the genera Dic-
tyotidium,Leiosphaeridia and Schizosporis could be
related to the green algae (Prasinophyceae and Zygne-
matophyceae). We follow this suggestion, without
providing arguments for or against this interpretation.
In addition, the vigourous debate of whether Reduvias-
poronites is of algal origin or a fungal spore continues
(Eshet et al. 1995; Visscher et al. 1996,2011; Afonin
et al. 2001; Foster et al. 2002; Sephton et al. 2009). We
do not discuss herein the biological affinity of Redu-
viasporonites, and tentatively place it in the acritarchs.
All acritarchs are classified as ‘Incertae sedis’, and listed
in alphabetical order.
Algae
Division Chlorophyta Pascher 1914
Class Prasinophyceae Christensen 1962
Order Not assigned
Family Pterosphaeridiaceae M€
adler 1963
Dictyotidium Eisenack 1955 emend. Staplin 1961
Type species. Dictyotidium dietyotum (Eisenack 1938)
Eisenack 1955
Dictyotidium reticulatum Schulz 1965
Plate 1, figures 1–12
Dictyotidium reticulatum Schulz 1965, p. 278, pl. 23,
figs. 12–14
Description. The vesicle is spherical to subspherical,
and covered by a reticulate ornamentation. The
ornamentation includes irregular and interconnected
fields, with four to six ridges. The width of each ridge is
ca. 4–10 mm. The wall is approximately 2–5 mm thick
Table 1. South Chinese species of the Micrhystridium/Very-
hachium complex recorded in the present study and their
attribution to five informal groups, based on their general
central body outline.
Group name
Shape of
the vesicle
Included in
this paper
V. cylindricum
group
ellipsoidal V. cylindricum
V. trispinosum
group
triangular V. hyalodermum V. sp.
V. laidii group rectangular V. nasicum V.
quadratum V.
sedecimspinosum
M. pentagonale
group
pentagonal &
hexagonal
M. pentagonale M.
stellatum
M. breve group spherical M. fragile M. breve
330 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
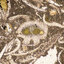
Plate 1. Phytoplankton from the latest Permian of South China. Each scale bar is 10 mm. Figures 1–9 were taken using SEM;
figures 10–12 were taken using the transmitted light microscope. Figures 1–12: Dictyotidium reticulatum. All specimens are from
Shangsi section/SS292-4.
Palynology 331
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
and usually folded. Folds are straight or slightly sinu-
ous. The wall occasionally splits open.
Dimensions. Vesicle diameter 52 (76) 110 mm, 45 speci-
mens measured.
Remarks. In Permian strata, the morphology and size
of Dictyotidium reticulatum are comparable to those of
Leiosphaeridia changxingesis Ouyang & Utting 1990.
However, the surface of the two species is very differ-
ent. The former is covered by an ornamentation of
irregular ridges, whereas the latter is nearly laevigate to
slightly scabrate, or punctuate with locally developed
ridges.
Previous records. Permian, China (Ouyang & Utting
1990); Norway (Mangerud 1994); Triassic, Germany
(Schulz 1965); Spain (Besems 1981).
Stratigraphical occurrence. Abundant in beds 15 to 22
of the Shangsi section, and present in the Xiakou
section.
Dictyotidium sp.
Plate 3, figures 11–12
Description. The vesicle is spherical. The surface of the
vesicle is covered by irregular and interconnected fields,
four to six-sided with ridges commonly 2–4 mm long,
the fields are mostly hollow.
Dimensions. Vesicle diameter 30 (36) 42 mm, five speci-
mens measured.
Remarks. The vesicle of this species is nearly spherical
in shape, and generally not folded. The fields are much
smaller than those of Dictyotidium reticulatum.
Stratigraphical occurrence. Present in bed 22 of the
Shangsi section.
Leiosphaeridia Eisenack 1958 emend. Downie &
Sarjeant 1963
Type species. Leiosphaeridia baltica Eisenack 1958
Leiosphaeridia microgranifera (Staplin 1961) Downie &
Sarjeant 1963
Plate 3, figures 19–21
Protoleiosphaeridium microgranifera Staplin 1961,
p. 405, pl. 48, fig. 4
Leiosphaeridia microgranifera (Staplin 1961) Downie &
Sarjeant 1963, p. 124
Description. The vesicle is ellipsoidal to spherical in
outline, commonly folded, with a dense granulose wall.
The granules are rounded, clearly separated, and about
0.5 mm in diameter.
Dimensions. Vesicle diameter 25 (32) 40 mm, 14 speci-
mens measured.
Remarks. This species is similar to Leiosphaeridia gran-
ulosa Staplin 1961. However, the vesicle of the former is
much larger and the granules are much smaller (0.5 mm
in diameter) than on the latter (1 mm diameter).
Previous records. Lower Silurian, Pennsylvania
(Johnson 1985); Upper Devonian, Canada (Staplin
1961).
Stratigraphical occurrence. Abundant in the Shangsi
and Xiakou sections.
Leiosphaeridia minutissima (Naumova 1949) Jankaus-
kas 1989
Plate 3, figures 13–18
Leiosphaeridia minutissima Naumova 1949, p. 52–53,
pl. 1, figs. 1–2, pl. 2, figs. 1–2
Leiosphaeridia minutissima (Naumova 1949) Jankaus-
kas 1989, p. 79–80, pl. 9, figs. 1–4, 11
Description. The vesicle is spherical to subspherical,
smooth, single-layered, always folded with the folds
being straight or slightly sinuous.
Dimensions. Vesicle diameter 25 (31) 40 mm, 23 speci-
mens measured.
Remarks. The outline of this species is similar to that of
Leiosphaeridia microgranifera, and both of them are
generally folded. However, the surfaces of the two spe-
cies are usually very different. L.minutissima has a
smooth vesicle wall, whereas L.microgranifera displays
dense granules.
Previous records. Neoproterozoic, Canada (Butterfield
& Chandler 1992); Congo (Gaucher & Germs 2006);
Czech Republic (Vavrdov
a2008); India (Bhat et al.
2009); Russia (Maslov 2004; Stanevich et al. 2007;
Vorob’eva et al. 2009); Ediacaran, Poland (Moczy-
dlowska 2008); Ukraine (Leonov & Ragozina 2007);
Uruguay (Gaucher et al. 2008); Cambrian, Czech
Republic (Konzalov
a1995; Steiner & Fatka 1996);
Russia (Naumova 1949)
Stratigraphical occurrence. Abundant in the Shangsi
and Dongpan sections.
Class Zygnematophyceae Round 1971
Order Zygnematales Borge & Pascher 1913
Family Zygnemataceae K€
utzing 1843
Schizosporis Cookson & Dettmann 1959 emend. Pierce
1976
Type species. Schizosporis reticulatus Cookson & Dett-
mann 1959
Schizosporis scissus (Balme & Hennelly 1956) Hart
1965
Plate 3, figures 8–10
Laevigatosporites scissus Balme & Hennelly 1956, p. 56,
pl. 1, figs. 6–9
Spheripollenites scissus (Balme & Hennelly 1956) Jan-
sonius 1962, p. 82, pl. 16, fig. 8
Schizosporis scissus (Balme & Hennelly 1956) Hart
1965,p.14
332 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Description. The vesicle is subspherical. The laevigate
wall is 0.5–1 mm thick, and occasionally splits open or
into two halves.
Dimensions. Vesicle diameter 25 (36) 45 mm, five speci-
mens measured.
Remarks. The wall of this species is much thicker than
that of other taxa, such as Micrhystridium,Very-
hachium,Leiosphaeridia and other genera, but it is
much smaller and thinner than that of Dictyotidium.
Generally, the vesicle splits into two halves and is not
folded.
Previous records. Permian, Antarctica (Balme &
Playford 1967; Farabee et al. 1991); Australia (Balme
& Hennelly 1956; Segroves 1967); Pakistan (Balme
1970); Triassic, Canada (Jansonius 1962).
Stratigraphical occurrence. Recorded from beds 22 and
25, Shangsi section.
Schizosporis sp.
Plate 3, figures 1–7
Description. The vesicle is spherical. Wall occasionally
splits open or into two halves. Dense short spines (ca.
0.5 mm long) present along the surface of the vesicle,
well distributed, simple, hollow and acuminate, closed
distally.
Dimensions. Vesicle diameter 20 (23) 25 mm, eight
specimens measured.
Remarks. Similar to Schizosporis scissus, the wall of
Schizosporis sp. splits open. Compared with Leiosphaeri-
dia microgranifera, it has a similar vesicle (wall thickness
and dense spines). However, whereas L. microgranifera
has a generally ellipsoidal and folded vesicle, that of
Schizosporis sp. is spherical and rarely folded.
Stratigraphical occurrence. Present in bed 25, Shangsi
section.
Group Acritarcha Evitt 1963
Gorgonisphaeridium Staplin et al. 1965
Type species. Gorgonisphaeridium winslowiae Staplin
et al. 1965
Gorgonisphaeridium sp.
Plate 2, figure 20
Description. The thin and single-layered vesicle is
spherical to subspherical, and covered by dense short
spines that are not pointed.
Dimensions. Vesicle diameter 25 (35) 42 mm, process
length 2–4 mm, four specimens measured.
Remarks. This species is very different from Micrhy-
stridium and Veryhachium, because of the spines and
the general outline. The vesicle is clearly much larger
than those of Micrhystridium, with dense spines on the
surface of the vesicle that are solid and not pointed.
Stratigraphical occurrence. Occurs in the Zhongzhai
and Shangsi sections.
Leiofusa Eisenack 1938
Type species. Leiofusa fusiformis Eisenack 1938
Leiofusa sp.
Plate 2, figure 13
Description. The central body is elliptical. The thin wall
is laevigate. Two long spines are present at both ends
of the central body.
Dimensions. Vesicle diameter 35 mm, spine length 17–
20 mm, one specimen measured.
Remarks. The central body is much bigger than that of
Micrhystridium and Veryhachium, but the spines are
similar. As only one specimen is recorded, it is left in
open nomenclature.
Stratigraphical occurrence. Present in bed 21, Shangsi
section.
Micrhystridium Deflandre 1937 emend. Sarjeant &
Stancliffe 1994
Type species. Micrhystridium inconspicuum (Deflandre
1935) Deflandre 1937
Micrhystridium breve Jansonius 1962
Plate 2, figures 21–25
Micrhystridium breve Jansonius 1962, p. 85, pl. 16, figs.
62, 63, 66
Description. The vesicle is ellipsoidal to spherical in
outline. The wall is single-layered and thin. About 20–
40 processes are present along the surface of the vesicle,
they are simple, hollow and acuminate, closed distally.
Vesicle and process surfaces are laevigate.
Dimensions. Vesicle diameter 15 (17) 21 mm, process
length 2–5 mm, 23 specimens measured.
Remarks. Sarjeant et al. (1970) suggested that Micrhy-
stridium breve and M.recurvatum forma brecispinosa
Valensi 1953 are possibly the same species. Because of
its numerous, short spines it is easily recognised.
Previous records. In Permian, Pakistan (Sarjeant et al.
1970); Canada (Utting 1978); Australia (McMinn
1982); USA (Jacobson et al. 1982); Brazil (Quadros
2002); In Triassic, Canada (Jansonius 1962; Utting
et al. 2004; Utting et al. 2005; Zonneveld et al. 2010);
Siberia (Ilyina & Egorov 2008); Cretaceous, Canada
(Collom & Hills 1999).
Stratigraphical occurrence. Abundant in beds 20 to 28,
Shangsi section.
Micrhystridium fragile Deflandre 1947
Plate 2, figure 18
Micrhystridium fragile Deflandre 1947, p. 8, fig. 13–18
Description. The vesicle is spherical to subspherical,
with a thin wall covered by tiny spines that are pointed
and generally flexible. Most of these spines are longer
than the diameter of the central body.
Palynology 333
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
334 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Dimensions. Vesicle diameter 9 (11) 12 mm, process
length 11–15 mm, four specimens measured.
Remarks. The vesicle of this species is the smallest in
our samples, only about 10 mm in diameter. The spines
are more flexible than those of other species of
Micrhystridium.
Previous records. Permian, Uruguay (Mautino et al.
1998); England (Sarjeant 1962); Triassic, England
(Van de Schootbrugge et al. 2007); Jurassic, England
(Wall 1965; Porter 1988; Stancliffe 1990); Israel (Sar-
jeant 1962).
Stratigraphical occurrence. Recorded from beds 21 to
25, Shangsi section.
Micrhystridium pentagonale Stockmans & Willi
ere
1963
Plate 2, figures 11–12
Micrhystridium pentagonale Stockmans & Willi
ere
1963, p. 470–471, pl. 3, fig. 32
Description. The thin-walled vesicle is pentagonal in
outline. Five prominent spines are present along each
corner. The other one to four spines arise from the cen-
tral area of the vesicle. All spines are pointed.
Dimensions. Vesicle diameter 12 (14) 17 mm, process
length 4–10 mm, 15 specimens measured.
Remarks. The outline of the vesicle is similar to that of
Micrhystridium stellatum, but it has a more pentagonal
shape. Moreover, the spines of the former are much
shorter.
Previous records. Carboniferous, Turkey (Higgs et al.
2002); Silurian, Belgium (Stockmans & Willi
ere 1963).
Stratigraphical occurrence. Present in beds 21 to 25 of
the Shangsi and Zhongzhai sections.
Micrhystridium stellatum Deflandre 1945
Plate 2, figure 19
Micrhystridium stellatum Deflandre 1945, p. 65, pl. 3,
figs. 16–19
Description. The vesicle is polyangular in outline. The
thin wall is single-layered. About eight to 12 strong but
simple processes are present around the vesicle, they
are relatively long, tapering, with sharp points and
hollow.
Dimensions. Vesicle diameter 13 (16) 18 mm, process
length 6–15 mm, 15 specimens measured.
Remarks. Common in Permian strata, the species can
easily be distinguished because of its polyangular vesi-
cle and the long spines.
Previous records. Ordovician, USA (Eley & Legault
1988); Silurian, England (Dorning 1981); France
(Deflandre 1945); Sweden (Gelsthorpe 2004; Stricanne
et al. 2006); Devonian, Libya (Moreau-Benoit 1984);
Poland (Filipiak 2009); Carboniferous, China (Gao
1986); Turkey (Higgs et al. 2002); Permian, Britain
(Wall & Downie 1963); China (Ouyang & Utting
1990); Germany (Schaarschmidt 1963); Israel (Horo-
witz 1973); Triassic, England (Van de Schootbrugge
et al. 2007).
Stratigraphical occurrence. Common in beds 21 to 28,
Shangsi section, and present in the Zhongzhai section.
Reduviasporonites Wilson 1962 emend. Foster et al.
2002
Type species. Reduviasporonites catenulatus Wilson
1962
Remarks. Reduviasporonites has been considered by
several authors to be a fungal spore. However, several
authors (e.g. Afonin et al. 2001; Foster et al. 2002)
questioned a fungal affinity, and suggested that it is
green algae. Because the biological affinity is unknown,
we place it here within the acritarchs. Interestingly,
many authors pointed out that the ‘spike’ of Reduvias-
poronites occurs at the PTB (e.g. Eshet et al. 1995;
Visscher et al. 1996). Indeed, around the PTB strata of
South China, the relative abundances of Reduviaspor-
onites are much higher than that in other strata, but
they never exceed 14% of an assemblage, which clearly
indicates that a ‘spike’ does not occur in the present
study.
Reduviasporonites catenulatus Wilson 1962
Plate 4, figures 12–17
Description. The vesicle is circular or oval in outline,
sometimes folded, wall laevigate. Many cells connect
together usually like a chain, but sometimes they are
present as pairs of cells or as a single cell. The length/
width ratio of the vesicle ranges between 1:1 and 2:1.
Plate 2. Phytoplankton from the latest Permian of South China. Each scale bar is 10 mm. All figures were taken using the trans-
mitted light microscope. 1–4: Veryhachium hyalodermum; 5–8: Veryhachium nasicum; 9–10: Veryhachium sedecimspinosum; 11–12:
Michrystridium pentagonale; 13: Leiofusa sp.; 14–15: Veryhachium cylindricum; 16: Veryhachium sp.; 17: Veryhachium quadratum;
18: Michrystridium fragile; 19: Michrystridium stellatum; 20: Baltisphaeridium sp.; 21–25: Michrystridium breve. The section/sam-
ple numbers for all specimens are as follows. 1–12: Shangsi section/SS25-E; 13: Shangsi section/SS289-1-2; 14–15, 17: Shangsi sec-
tion/SS290-7; 16: Shangsi section/SS291-6; 18: Shangsi section/SS290-4; 19, 21–25: Shangsi section/TS28-18; 20: Zhongzhai
section/ZZ22-2.
J
Palynology 335
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
336 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Dimensions. Vesicle diameter 8 (15) 20 mm, 17 speci-
mens measured.
Remarks. Foster et al. (2002) described this species in
detail, indicating that many characteristics are shared
between Reduviasporonites catenulatus and Reduvias-
poronites chalastus. We simply distinguish them on the
basis that the former is much smaller than the latter.
Previous records. Permian, Australia (Foster et al.
2002); USA (Wilson 1962; Elsik 1999); Cretaceous,
Canada (Kalgutkar & Braman 2008).
Stratigraphical occurrence. Common in the Shangsi
section, and present in the Xiakou section.
Reduviasporonites chalastus (Foster 1979) Elsik 1999
Plate 4, figures 1–11, 18–23
Chordecystia chalasta Foster 1979, p. 109–110, pl. 41,
figs. 3–9
?Brazilea helbyi forma gregata Foster 1979, p. 112, pl.
41, figs. 1–2
Tympanicysta stoschiana Balme 1979, p. 22–24, pl. 1,
figs. 3–7
Tympanicysta stoschiana Afonin et al. 2001, p. 484–
486, figs. 1, 2A–C, E, F
Reduviasporonites stoschianus (Balme 1979) Elsik 1999,
p. 40, pl. 1, figs. 1–24.
?Reduviasporonites stoschianus (Balme 1979) Elsik
1999, Wood & Elsik 1999, p. 46–48, pl. 1, figs. 1–9, pl.
2, figs. 1–7
Description. The vesicle is subcircular or subrectangu-
lar, sometimes folded, smooth. Many cells connect
together, usually forming a chain, but sometimes they
are present as pairs of cells or isolated as a single cell.
The length/width ratio of the vesicle varies between 2:1
and 6:1.
Dimensions. Vesicle diameter 15 (36) 65 mm, 19 speci-
mens measured.
Remarks. The species is distinguished based on the
characteristic chain-forming cells, the vesicle being
much larger than that of Reduviasporonites catenulatus.
Previous records. Permian, Australia (Foster 1979; Fos-
ter et al. 2002); China (Ouyang & Utting 1990); Den-
mark (Balme 1979); Iraq (Stolle 2007); Kenya (Hankel
1992); Paraguay (P
erez Loinaze et al. 2010); Peru
(Wood & Elsik 1999); Russia (Afonin et al. 2001); Tur-
key (Stolle 2010); USA (Elsik 1999).
Stratigraphical occurrence. Common in the Shangsi
and Zhongzai sections.
Veryhachium Deunff 1954 emend. Sarjeant & Stancliffe
1994
Type species. Veryhachium trisulcum (Deunff 1954)
Deunff 1959
Veryhachium cylindricum Schaarschmidt 1963
Plate 2, figures 14, 15
Veryhachium cylindricum Schaarschmidt 1963,p.64,
pl. 18, figs. 8–10
Description. The vesicle is ellipsoidal in outline. The
thin wall is single-layered. Six prominent but simple
processes are present around the vesicle, they are proxi-
mally wide (about 5 mm in width), and the tips are acu-
minate, and closed distally.
Dimensions. Vesicle diameter 15 (18) 22 mm, process
length 8–12 mm, 13 specimens measured.
Remarks. This species is easily recognisable, because of
its six spines that are arranged symmetrically around
the vesicle. Two of them prolong the long axis of the
vesicle, and the other four spines are displayed sym-
metrically around the short axis of the vesicle.
Previous records. Permian, Germany (Schaarschmidt
1963).
Stratigraphical occurrence. Recorded from beds 21 to
25, Shangsi section.
Veryhachium hyalodermum (Cookson 1956) Schaarsch-
midt 1963
Plate 2, figures 1–4
Veryhachium hyalodermum Cookson 1956, p. 188–189,
pl. 1, figs. 12–16
Veryhachium hyalodermum (Cookson 1956) Schaarsch-
midt 1963, p. 62–63
Description. The vesicle is triangular in outline, with
four processes. Three processes are present along each
angle of the vesicle, and another process is attached on
the central body. The processes are simple, long, and
distally closed. The thin wall is single layered.
Dimensions. Vesicle diameter 10 (12) 14 mm, process
length 9–15 mm, 25 specimens measured.
Remarks. Many species of Veryhachium with four
spines or a triangular vesicle have been described, such
as V.ceratioides Stockmans & Willi
ere 1962,
Plate 3. Phytoplankton from the latest Permian of South China. Each scale bar is 10 mm. Figures 11–12 were taken using scan-
ning electron microscopy (SEM); figures 1–10 and 13–21 were taken using the transmitted light microscope.1–7: Schizosporis sp.;
8–10: Schizosporis scissus; 11–12: Dictyotidium sp.; 13–18: Leiosphaeridia minutissima; 19–21: Leiosphaeridia microgranifera. The
section/sample numbers for all specimens are as follows. 1–10: Shangsi section/SS25-E; 11–12: Shangsi section/SS292-4; 13–14:
Dongpan section/DP2-G; 15–16: Dongpan section/DP3-A; 17: Dongpan section/DP5-B; 18: Shangsi section/SS290-8-1; 19–21:
Shangsi section/SS291-2.
J
Palynology 337
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
338 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
V.europaeum Stockmans & Willi
ere 1960,V.leonense
Cramer 1964 and others. It is difficult to distinguish
V.hyalodermum and V.europaeum solely by their
vesicle outline. However, the spines of the former are
obviously much longer than those of the latter.
Previous records. Permian, China (Ouyang & Utting
1990); Germany (Schaarschmidt 1963); Israel (Horo-
witz 1974); South Africa (Horowitz 1990); Eocene,
Australia (Cookson 1956).
Stratigraphical occurrence. Common in beds 21 to 28,
Shangsi section.
Veryhachium nasicum (Stockmans & Willi
ere 1960)
Stockmans & Willi
ere 1962
Plate 2, figures 5–8
Stellinium nasicum Stockmans & Willi
ere 1960,p.3,pl.
1, fig. 3
Veryhachium nasicum (Stockmans & Willi
ere 1960)
Stockmans & Willi
ere 1962,p.52
Description. The vesicle is rectangular in outline, with
five processes. Four of these processes are present at
each corner of the vesicle, and another process arises
from the central body; the processes are simple, short,
and distally closed. The thin wall is single layered.
Dimensions. Vesicle diameter 17 (19) 22 mm, process
length 5–9 mm, 21 specimens measured.
Remarks. There are many species of Veryhachium with
a rectangular vesicle and Servais et al. (2007) attrib-
uted them to the Veryhachium lairdii group. V. nasicum
is easily recognised with its characteristic four spines
present at each corner, and a further spine attached on
the surface of the vesicle.
Previous records. Devonian, Belgium (Stockmans &
Willi
ere 1960); China (Gao 1986); Libya (Moreau-
Benoit 1984); Carboniferous, Russia (Marhoumi &
Rauscher 1984); Permian, Germany (Schaarschmidt
1963).
Stratigraphical occurrence. Common in beds 21 to 25,
Shangsi section.
Veryhachium quadratum Schaarschmidt 1963
Plate 2, figure 17
Veryhachium quadratum Schaarschmidt 1963, p. 63, pl.
17, figs. 8–10
Description. The vesicle is polyangular in outline. The
thin wall is single-layered, with about five to eight short
spines that are 2 mm wide at their base, and 1–2 mm
long.
Dimensions. Vesicle diameter about 20 mm, process
length 1–2 mm, one specimen measured.
Remarks. This species is very different from the other
species of Veryhachium, because of its short spines,
generally only 1–2 mm in length.
Previous records. Permian, Germany (Schaarschmidt
1963).
Stratigraphical occurrence. Present in bed 21, Shangsi
section.
Veryhachium sedecimspinosum Staplin 1961
Plate 2, figures 9–10
Veryhachium sedecimspinosum Staplin 1961, p. 414, pl.
49, figs. 9–11
Description. The thin wall is single-layered, and the
outline is rectangular, with five to eight processes.
Four of these processes arise from each corner of the
vesicle, and the other processes (two to four in number)
arise from the central body. All of the processes are
simple, short, and distally closed.
Dimensions. Vesicle diameter 15 (16) 17 mm, process
length 7–13 mm, four specimens measured.
Remarks. Similar to Veryhachium nasicum, the rectan-
gular vesicle of V. sedecimspinosum has four spines that
arise from each corner. However, there are more (gen-
erally two to four) additional spines attached on the
surface of the vesicle.
Previous records. Devonian, Canada (Staplin 1961);
Permian, Germany (Schaarschmidt 1963).
Stratigraphical occurrence. Common in the Shangsi
section.
Veryhachium sp.
Plate 2, figure 16
Description. The vesicle is triangular with a smooth
wall. Three short spines are present at each of the
angles. One or two short additional spines arise from
the central body.
Dimensions. Vesicle diameter 25 mm, process length
about 5 mm, one specimen measured.
Remarks. Similar to Veryhachium hyalodermum,Very-
hachium sp. has a triangular vesicle, but it is much
larger than the latter, and the spines are much shorter.
Because only one specimen has been recovered, the
species is left in open nomenclature.
Stratigraphical occurrence. Present in bed 21, Shangsi
section.
Plate 4. Phytoplankton from the latest Permian of South China. Each scale bar is 10 mm. All figure images were taken using the
transmitted light microscope. 1–11, 18–23: Reduviasporonites chalastus; 12–17: Reduviasporonites catenulatus. The section/sample
numbers for all specimens are as follows. 1–6, 11, 14, 23: Shangsi section/TS28-17; 12–13: Xiakou section/XK254-1; 16–17:
Zhongzhai section/ZZ19-3; 7–10, 15, 18–22: Shangsi section/ TS28-18.
J
Palynology 339
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Acknowledgements
We express sincere thanks to Hu Qing, Zhang Lei, Liu
Guichun, Shen Jun, Xiang Yu, and Miss Xu Guozhen and
Nie Xiaomei for their help with the fieldwork. We acknowl-
edge the two referees (Reed Wicander, Central Michigan
University, USA, and Clinton Foster, Geoscience Australia)
for their detailed reviews and helpful comments, and the
Editor of the journal, Jim Riding (British Geological
Survey), for his help and advice. This work was supported by
National Science Foundation of China (NSFC) Project num-
bers 40839903, 40921062, the Chinese 973 (2011CB808800),
and 111 programmes (B08030), the Ministry of Education of
China (NCET-10-0712), the Fundamental Funds for the
Central Universities (CUGL100504), the Foundation of the
Geological Survey of China (1212011220529), and by grants
of the University of Lille1 (‘Allocation Pr
esidentielle’ and
‘Bonus Qualit
e Recherche’) and the Conseil R
egional du
Nord (France).
Author biographies
YONG LEI is a Ph.D. Candidate at
the China University of Geosciences,
Wuhan and the University of Lille 1,
France. He is to receive his Ph.D.
during 2013; the thesis focuses on the
diversity and ecology of acritarchs
around the Permian-Triassic boundary
in southern China. His research inter-
ests include Permian palynology and
palynofacies, and the end-Permian extinction event.
THOMAS SERVAIS is a research
director at the French Centre of Scien-
tifical Research (CNRS). He studied
geology at the universities of Namur
and Li
ege in Belgium, and received a
Ph.D. on Ordovician acritarchs in 1993.
After post-doctoral studies in Belgium,
Germany and the United Kingdom,
Thomas was recruited as a CNRS
research associate at the University of Lille 1 in 1997. Most
of his research is on Lower Palaeozoic microphytoplankton,
but other fields of interest include regional geology, macropa-
laeontology (corals, graptolites and trilobites) and calcareous
micropalaeontology (e.g. calcareous microfossils). Thomas is
currently Past-President of the International Federation of
Palynological Societies (IFPS), and Vice-President of the
Palaeontological Association.
QINLAI FENG is a geology professor
at China University of Geosciences,
Wuhan. His research interests include
the Permian-Triassic mass extinction
event, Palaeozoic and Triassic radiolar-
ians, and the palaeontology of south-
west China and southeast Asia.
WEIHONG HE is a professor of
Geosciences at the China University of
Geosciences, Wuhan. She has a Ph.D.
in palaeontology and stratigraphy from
the China Institute of Geosciences,
Beijing, which was awarded in 2002.
Her research interests include Permian-
Triassic bivalves, brachiopods, radio-
larians and other invertebrates, mass
extinctions, palaeogeography and palaeoecology.
References
Afonin SA, Barinova SS, Krassilov VA. 2001. A bloom of
Tympanicysta Balme (green algae of zygnematalean affin-
ities) at the Permian-Triassic boundary. Geodiversitas 23
(4):481–487.
Balme BE. 1970. Palynology of Permian and Triassic strata
in the Salt range and Surghar range, West Pakistan.
Special Publication, University of Kansas, Department
of Geology 4:305–453.
Balme BE. 1979. Palynology of Permian-Triassic boundary
beds at Kap Stosch, East Greenland. Medd Grønland
200(6):1–37.
Balme BE, Hennelly JPF. 1956. Monolete, monocolpate, and
alete sporomorphs from Australian Permian sediments.
Aust J Bot. 4(1):54–67.
Balme BE, Playford G. 1967. Late Permian plant microfossils
from the Prince Charles Mountains, Antarctica. Rev
Micropal
eontol. 10(3):179–192.
Besems RE. 1981. Aspects of middle and late Triassic paly-
nology. 2. Preliminary palynological data from the Hor-
nos-Siles Formation of the Prebetic Zone, NE province
of Jaen (Southeastern Spain). Rev Palaeobot Palynol. 32
(4):389–400.
Bhat GM, Ram G, Koul S. 2009. Potential for oil and gas in
the Proterozoic carbonates (Sirban Limestone) of
Jammu, northern India. Geol Soc Lond Special Pub. 326
(1):245–254.
Borge O, Pascher A. 1913. Zygnemales. In: Pascher A, editor.
Die S€
ußwasserflora Deutschlands,
€
Osterreichs und der
Schweiz. Jena (Germany): Gustav Fischer Verlag; p. 9, 51.
Butterfield NJ, Chandler FW. 1992. Paleoenvironmental dis-
tribution of Proterozoic microfossils, with an example
from the Agu Bay Formation, Baffin Island. Palaeontol-
ogy 35(4):943–957.
Christensen K. 1962. Alger. In: Bocher TW, Lange M,
Sorensen T, editors. Botanik, 2. Copenhagen: Munks-
gaard; p. 178.
Colbath GK, Grenfell HR. 1995. Review of biological affini-
ties of Paleozoic acid-resistant, organic-walled eukaryotic
algal microfossils (including “acritarchs”). Rev Palaeobot
Palynol. 86(3–4):287–314.
Collom CJ, Hills LV. 1999. Geological Survey of Canada,
Open File 3833. Victoria (British Columbia): Natural
Resources Canada Ottawa; p. 37.
Cookson IC. 1956. Additional Microplankton from Austra-
lian Late Mesozoic and Tertiary Sediments. Mar Fresh-
wat Res. 7(1):183–191.
Cookson IC, Dettmann ME. 1959. On Schizosporis, a new
form genus from Australian Cretaceous deposits. Micro-
paleontology 5(2):213–216.
340 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Cramer FH. 1964. Microplankton from three Palaeozoic
formations in the Province of Le
on, NW Spain. Leidse
Geol Med. 30:253–361.
Deflandre G. 1935. Consid
erations biologiques sur les micro-
organismes d’origine planctonique conserv
es dans les
silex de la craie. Bull Biol France Belg. 69:213–244.
Deflandre G. 1937. Microfossiles des silex cr
etac
es:
Flagell
es incertae sedis, Hystrichosphaerid
es, Sarcodin
es,
organismes divers. Annal Pal
eontol. 26:51–103.
Deflandre G. 1945. Microfossiles des calcaires siluriens de la
Montagne Noire. Annal Pal
eontol. 31:41–75.
Deflandre G. 1947. Le probl
eme des Hystrichosph
eres. Bull
l’Institut Oc
eanograph (Monaco) 919: 23.
Deunff J. 1954. Veryhachium, genre nouveau d’Hystri-
chosph
eres du Primaire. Comp Rendu Somm S
eances
Soci G
eol France 13:305–306.
Deunff J. 1959. Microorganismes planctoniques du Primaire
armoricain: Ordovicien du Veryhac’h (presqu’^
ıle de Cro-
zon). Bull Soc G
eol min
eral Bretagne 2:1–41.
Dorning KJ. 1981. Silurian acritarchs from the type Wenlock
and Ludlow of Shropshire, England. Rev Palaeobot
Palynol. 34(2):175–203.
Downie C, Sarjeant WAS. 1963. On the interpretation and
status of some hystrichosphere genera. Palaeontology 6
(1):83–96.
Eisenack A. 1938. Hystrichosph€
aerideen und verwandte For-
men im baltischen Silur. Zeits Geschiebeforschung
Flachlandgeol. 14(1):1–30.
Eisenack A. 1955. Chitinozoen, Hystrichosph€
aren und
andere Mikrofossilien aus dem Beyrichia-Kalk. Sencken-
bergiana Lethaea 36(1–2):157–188.
Eisenack A. 1958. Mikroplankton aus dem norddeutschen
Apt, nebst einigen Bemerkungen €
uber fossile Dinoflagel-
laten. Neues Jahr Geol Pal€
aontol Abhand. 39(5–6):389–
405.
Eisenack A. 1969. Zur systematik einiger pal€
aozoischer Hys-
trichosph€
aren (Acritarcha) des baltischen Gebietes.
Neues Jahr Geol Pal€
aontol Abhand. 133(3):245–266.
Eley BE, Legault JA. 1988. Palynomorphs from the Mani-
toulin Formation (Early Llandovery) of southern
Ontario. Palynology 12(1):49–63.
Elsik WC. 1999. Reduviasporonites Wilson 1962: Synonymy
of the fungal organism involved in the late Permian crisis.
Palynology 23:37–41.
Eshet Y, Rampino MR, Visscher H. 1995. Fungal event and
palynological record of ecological crisis and recovery
across the Permian-Triassic boundary. Geology 23
(11):967–970.
Evitt WR. 1963. A discussion and proposals concerning fossil
dinoflagellates, hystrichospheres, and acritarchs, II. Proc
Nat Acad Sci USA 49(3):298–302.
Farabee MJ, Taylor EL, Taylor TN. 1991. Late Permian
palynomorphs from the Buckley Formation, central
Transantarctic Mountains, Antarctica. Rev Palaeobot
Palynol. 69(4):353–368.
Feng QL, Gu SZ. 2002. Uppermost Changxingian (Permian)
radiolarian fauna from southern Guizhou, southwestern
China. J Paleontol. 76(5):797–809.
Feng QL, He WH, Gu SZ, Meng YY, Jin YX, Zhang F.
2007. Radiolarian evolution during the latest Permian in
South China. Global Planet Change 55(1–3):177–192.
Fensome RA, Williams GL, Barss MS, Freeman JM, Hill JM.
1990. Acritarchs and fossil Prasinophytes: An index to
genera, species and intraspecific taxa. American Associa-
tion of Stratigraphic Palynologists Foundation 25;771 p.
Filipiak P. 2009. Lower Famennian phytoplankton from the
Holy Cross Mountains, Central Poland. Rev Palaeobot
Palynol. 157(3–4):326–338.
Foster CB. 1979. Permian plant microfossils of the Blair
Athol Coal Measures, Baralaba Coal Measures, and
basal Rewan Formation of Queensland. Geological Sur-
vey of Queensland Publications (372); 244 p.
Foster CB, Stephenson MH, Marshall C, Logan GA, Green-
wood PF. 2002. A revision of Reduviasporonites Wilson
1962: description, illustration, comparison and biological
affinities. Palynology 26(1):35–58.
Gao LD. 1986. Late Devonian and Early Carboniferous acri-
tarchs from Nyalam county, Xizang (Tibet), China. Rev
Palaeobot Palynol. 47(1–2):17–30.
Gaucher C, Blanco G, Chiglino, L, Poir
e D, Germs GJB.
2008. Acritarchs of Las Ventanas Formation (Ediacaran,
Uruguay): Implications for the timing of coeval rifting
and glacial events in western Gondwana. Gondwana
Res. 13(4):488–501.
Gaucher C, Germs GJB. 2006. Recent advances in South
African Neoproterozoic-Early Palaeozoic biostratigra-
phy: correlation of the Cango Caves and Gamtoos
Groups and acritarchs of the Sardinia Bay Formation,
Saldania Belt. S Afr J Geol. 109(1–2):193–214.
Gelsthorpe DN. 2004. Microplankton changes through the
early Silurian Ireviken extinction event on Gotland, Swe-
den. Rev Palaeobot Palynol. 130(1–4):89–103.
Grice K, Twitchett RJ, Alexander R, Foster CB, Looy CV.
2005. A potential biomarker for the Permian–Triassic
ecological crisis. Earth Planet Sci Lett. 236(1–2):315–321.
Hankel O. 1992. Late Permian to early Triassic microfloral
assemblages from the Maji y a Chumvi Formation,
Kenya. Rev Palaeobot Palynol. 72(1–2):129–147.
Hart GF. 1965. The systematics and distribution of Permian
miospores. Johannesburg, South Africa: Witwatersrand
University Press; p. 252.
Hays LE, Grice K, Foster CB, Summons RE. 2011. Bio-
marker and isotopic trends in a Permian–Triassic sedi-
mentary section at Kap Stosch, Greenland. Org
Geochem. 43:67–82.
He WH, Feng QL, Weldon EA, Gu SZ, Meng YY, Zhang F.,
Wu S.B. 2007a. A late Permian to early Triassic bivalve
fauna from the Dongpan Section, southern Guangxi,
South China. J Paleontol. 81(5):1009–1019.
He WH, Shen SZ, Feng QL, Gu SZ. 2005. A late Changhsin-
gian (Late Permian) deepwater brachiopod fauna from
the Talung Formation at the Dongpan section, southern
Guangxi, South China. J Paleontol. 79(5):927–938.
He WH, Shi GR, Feng QL, Campi MJ, Gu SZ, Bu JZ, Peng
YQ, Meng YY. 2007b. Brachiopod miniaturization and
its possible causes during the Permian–Triassic crisis in
deep water environments, South China. Palaeogeogr
Palaeoclimatol Palaeoecol. 252(1):145–163.
He WH, Shi GR, Gao YQ, Peng YQ, Zhang Y. 2008. A new
Early Triassic microgastropod fauna from the Zhongzhai
Section, Guizhou, southwestern China. Proc Royal Soc
Vic. 120(1):157–166.
Higgs KT, Finucane D, Tunbridge IP. 2002. Late Devonian
and early Carboniferous microfloras from the HakkarI
Province of southeastern Turkey. Rev Palaeobot Palynol.
118(1–4):141–156.
Palynology 341
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Hong HL, Xie SC, Lai XL. 2011. Volcanism in association
with the prelude to mass extinction and environment
change across the Permian-Triassic boundary (PTB),
southern China. Clay Miner. 59(5):478–489.
Hong HL, Zhang N, Li ZH, Xue H, Xia W, Yu N. 2008.
Clay mineralogy across the PT boundary of the Xiakou
section, China: Evidence of clay provenance and environ-
ment. Clay Miner. 56(2):131–143.
Horowitz A. 1973. Late Permian palynomorphs from south-
ern Israel. Pollen Spore. 15(2):315–341.
Horowitz A. 1974. Esp
eces du genre Veryhachium du Permo-
Trias du Sud d’Israel. Rev Micropal
eontol. 17(2):75–80.
Horowitz A. 1990. Palynology and paleoenvironment of ura-
nium deposits in the Permian Beaufort Group, South
Africa. Ore Geol Rev. 5(5–6):537–540.
Ilyina NV, Egorov AY. 2008. The Upper Triassic of northern
Middle Siberia: stratigraphy and palynology. Polar Res.
27(3):372–392.
Jacobson SR, Wardlaw BR, Saxton JD. 1982. Acritarchs from
the Phosphoria and Park City Formations (Permian,
Northeastern Utah). J Paleontol. 56(2):449–458.
Jankauskas TV. 1989. Mikrofossilii dokembriia SSSR. Len-
ingrad: Nauka; 188 p.
Jansonius J. 1962. Palynology of Permian and Triassic sedi-
ments, Peace River area, western Canada. Palaeonto-
graph Abt B 110(1–4):35–98.
Jiang HS, Lai XL, Yan CB, Aldridge R, Wignall PB, Sun
YD. 2011. Revised conodont zonation and conodont
evolution across the Permian–Triassic boundary at the
Shangsi section, Guangyuan, Sichuan, South China.
Global Planet Change 77(3–4):103–115.
Johnson NG. 1985. Early Silurian palynomorphs from the
Tuscarora Formation in central Pennsylvania and their
paleobotanical and geological significance. Rev Palaeo-
bot Palynol. 45(3–4):307–359.
Kalgutkar RM, Braman DR. 2008. Santonian To ?Earliest
Campanian (Late Cretaceous) Fungi from the Milk River
Formation, Southern Alberta, Canada. Palynology 32
(1):39.
Konzalov
a OFM. 1995. Microfossils of the Paseky Shale
(Lower Cambrian, Czech Republic). J Czech Geol Soc.
40(4):55–66.
K€
utzing FT. 1843. Phycologia generalis. Leipzig: Brockhaus;
p. 458.
Lai XL, Yang FQ, Hallam A, Wignall P. 1996. The Shangsi
section, candidate of the Global Stratotype Section and
Point of the Permian-Triassic boundary. In: Yin HF, edi-
tor. The Palaeozoic-Mesozoic Boundary. Candidates of
Global Stratotype Section and Point of the Permian-
Triassic Boundary. Wuhan: China University of Geo-
sciences Press; p. 113–124.
Le H
eriss
e A, Dorning KJ, Mullins GL, Wicander R. 2009.
Global patterns of organic-walled phytoplankton biodi-
versity during the Late Silurian to Earliest Devonian.
Palynology 33(1):25–75.
Lei Y, Servais T, Feng QL. 2013. The diversity of the Perm-
ian phytoplankton. Rev Palaeobot Palynol. doi: http://
dx.doi.org/10.1016/j.revpalbo.2013.03.004.
Lei Y, Servais T, Feng QL, He WH. 2012. The spatial (near-
shore-offshore) distribution of latest Permian phytoplank-
ton from the Yangtze Block, South China. Palaeogeogr
Palaeoclimatol Palaeoecol. 363–364:151–162.
Leonov MV, Ragozina AL. 2007. Upper Vendian assemb-
lages of carbonaceous micro-and macrofossils in the
White Sea Region: systematic and biostratigraphic
aspects. Geol Soc Lond Special Pub. 286(1):269–275.
Li J, Servais T, Brocke R. 2002. Chinese Paleozoic acritarch
research: review and perspectives. Rev Palaeobot
Palynol. 118(1–4):181–193.
Li J, Cao CQ, Servais T, Zhu YH. 2004. Later Permian acri-
tarchs from Meishan (SE China) in the context of Perm-
ian palaeobiogeography and palaeoecology. Neues Jahrb
Geol Pal€
aontol Monatshefte 2004–7:427–448.
Li ZS, Zhan LP, Zhu XF, Zhang JH, Jin RG, Liu GF, Sheng
HB, Shen GM, Dai JY, Huang HQ, Xie LC, Yan Z.
1986. Mass extinction and geological events between
Palaeozoic and Mesozoic era. Acta Geol Sin. 60:1–15.
M€
adler K. 1963. Die figurierten organischen Bestandteile der
Posidonienschiefer. Geologisch Jahr Beihefte 58:287–406.
Mangerud G. 1994. Palynostratigraphy of the Permian
and lowermost Triassic succession, Finnmark Platform,
Barents Sea. Rev Palaeobot Palynol. 82(3–4):317–349.
Marhoumi R, Rauscher R. 1984. Un plancton D
evonien de
la M
eseta orientale au Maroc. Rev Palaeobot Palynol. 43
(1–3):237–253.
Maslov AV. 2004. Riphean and Vendian sedimentary
sequences of the Timanides and Uralides, the eastern
periphery of the East European Craton. Geol Soc Lond
Mem. 30(1):19–35.
Mautino LR, Vergel MDM, Anz
otegui LM. 1998. Palinolo-
gia de la Formacion Melo (Permico inferior) en Arroyo
Seco, Departamento Rivera, Uruguay, Parte V: Granos
de polen, acritarcas E Incertae sedis. Ameghiniana 35
(3):299–314.
McMinn A. 1982. Late Permian acritarchs from the northern
Sydney Basin. J Proc Royal Soc NSW 115:79–86.
Metcalfe I, Nicoll RS. 2007. Conodont biostratigraphic con-
trol on transitional marine to non-marine Permian-
Triassic boundary sequences in Yunnan-Guizhou, China.
Palaeogeograph Palaeoclimatol Palaeoecol. 252(1–2):56–
65.
Moczyd»owska M. 2008. The Ediacaran microbiota and the
survival of Snowball Earth conditions. Precamb Res. 167
(1–2):1–15.
Moreau-Benoit A. 1984. Acritarches et chitinozoaires du
D
evonien moyen et sup
erieur de Libye occidentale. Rev
Palaeobot Palynol. 43(1–3):187–216.
Mullins GL, Servais T. 2008. The diversity of the Carbonifer-
ous phytoplankton. Rev Palaeobot Palynol. 149(1–2):29–
49.
Naumova SN. 1949. Spores of the Lower Cambrian. Izves-
tiya Akad Nauk SSSR, Ser Geolog. 4:49–56.
Ouyang S. 1982. Upper Permian and Lower Triassic palyno-
morphs from eastern Yunnan, China. Can J Earth Sci. 19
(1):68–80.
Ouyang S. 1986. Palynology of Upper Permian and Lower
Triassic strata of Fuyuan district, Eastern Yunan. Palae-
ont Sin. 169(9):1–122.
Ouyang S, Utting J. 1990. Palynology of upper Permian and
lower Triassic rocks, Meishan, Changxing County,
Zhejiang Province, China. Rev Palaeobot Palynol.
66(1–2):65–103.
Pascher A. 1914.
€
Uber Flagellaten und Algen. Berichte
Deutsch Botanisch Gesellschaft 32:136–160.
P
erez Loinaze VS, C
esari SN, L
opez Gamund
ı O, Buatois L.
2010. Palynology of the Permian San Miguel Formation
(Western Paran
a Basin, Paraguay): Gondwanan bio-
stratigraphic correlations. Geol Acta 8(4):483–493.
342 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
Pierce ST. 1976. Morphology of Schizosporis reticulatus Cook-
son and Dettmann 1959. Geoscience Man 15(1):25–33.
Porter R. 1988. Palynological evidence for Jurassic
microplankton provinces in Great Britain. Rev Palaeobot
Palynol. 56(1–2):21–39.
Quadros LP. 2002. Acritarcos e tasmanites do Permo-
Carbon
ıfero da bacia do Paran
a. Rev Inst Geol. 23
(1):39–50.
Riegel W. 1996. The geologic significance of the Late Paleozoic
phytoplankton blackout. IX IPC Meeting. Abstracts
volume. Houston (TX): American Association of Strati-
graphic Palynologists; 133–134.
Riegel W. 2008. The Late Palaeozoic phytoplankton black-
out - Artefact or evidence of global change. Rev Palaeo-
bot Palynol. 148(2–4):73–90.
Round FE. 1971. The taxonomy of the Chlorophyta. II.
British Phycolog J. 6(2):235–264.
Sarjeant WAS. 1962. Microplankton from the Ampthill
Clay of Melton, South Yorkshire. Palaeontology
5(3):478–497.
Sarjeant WAS, Kummel B, Teichert C. 1970. Acritarchs and
tasmanitids from the Chhidru Formation, uppermost
Permian of West Pakistan. Stratigraphical Boundary
Problems: Permian and Triassic of West Pakistan. Univ
Kansas Special Pub. 4:277–304.
Sarjeant WAS, Stancliffe RPW. 1994. The Micrhystridium
and Veryhachium complexes (Acritarcha: Acanthomor-
phitae and Polygonomorphitae); a taxonomic reconsider-
ation. Micropaleontology 40(1):1–77.
Schaarschmidt F. 1963. Sporen und Hystrichosphaerideen
aus dem Zechstein von B€
udingen in der Wetterau. Palae-
ontograp Abt B 113(1–4):38–91.
Schulz E. 1965. Sporae dispersae aus der Trias von Th€
urin-
gen. Abhandlungen des ZGI. 1:257–287.
Segroves KL. 1967. Cutinized microfossils of probable non-
vascular origin from the Permian of Western Australia.
Micropaleontology 13(3):289–305.
Sephton MA, Visscher H, Looy CV, Verchovsky AB,
Watson JS. 2009. Chemical constitution of a Permian-
Triassic disaster species. Geology 37(10):875–878.
Servais T. 1996. Some considerations on acritarch classifica-
tion. Rev Palaeobot Palynol. 93(1–4):9–22.
Servais T, Lehnert O, Li J, Mullins GL, Munnecke A, N€
utzel
A, Vecoli M. 2008. The Ordovician Biodiversification: rev-
olution in the oceanic trophic chain. Lethaia 41(2):99–109.
Servais T, N€
utzel A, Mullins GL. 2006. Was there a phyto-
plankton blackout in the late Paleozoic? Palynology
30:228.
Servais T, Li J, Stricanne L, Vecoli M, Wicander R. 2004.
Acritarchs. In: Webby BD, Droser ML, Paris F, Percival
IG, editors. The Great Ordovician Biodiversification
Event. New York: Columbia University Press; p. 348–360.
Servais T, Vecoli M, Li J, Molyneux SG, Raevskaya EG,
Rubinstein C, Claudia V. 2007. The acritarch genus Ver-
yhachium Deunff 1954: taxonomic evaluation and first
appearance. Palynology 31:191–203.
Shen J, Algeo TJ, Hu Q, Zhang N, Zhou L, Xia WC, Xie SC,
Feng QL. 2012a. Negative C-isotope excursions at the
Permian-Triassic boundary linked to volcanism. Geology
40:963–966.
Shen J, Algeo TJ, Zhou L, Feng QL, Yu JX, Ellwood B.
2012b. Volcanic perturbations of the marine environ-
ment in South China preceding the latest Permian mass
extinction and their biotic effects. Geobiology 10(1):82–
103.
Shen SZ, Crowley JL, Wang Y, Bowring SA, Erwin DH,
Sadler PM, Cao CQ, Rothman DH, Henderson CM,
Ramezani J. 2011. Calibrating the end-Permian mass
extinction. Science 334(6061):1367–1372.
Stancliffe RPW. 1990. Acritarchs and other non-
Dinophycean marine palynomorphs from the Oxfordian
(Upper Jurassic) of Skye, Western Scotland and Dorset,
Southern England. Palynology 14:175–192.
Stanevich AM, Maksimova EN, Kornilova TA, Mazukab-
zov AM, Gladkochub DP. 2007. Microfossils of the late
Proterozoic Debengdinskaya Formation of the Olenek-
skiy uplift. Bull Tomsk Polytechnic Univ. 311(1):9–14.
Staplin FL. 1961. Reef-controlled distribution of Devonian
microplankton in Alberta. Palaeontology 4(3):392–424.
Staplin FL, Jansonius J, Pocock SAJ. 1965. Evaluation of
some acritarchous hystrichosphere genera. Neues Jahr
Geol Pal€
aontol Abhand. 123(2):167–201.
Steiner M, Fatka O. 1996. Lower Cambrian tubular micro-to
macrofossils from the Paseky Shale of the Barrandian
area (Czech Republic). Pal€
antol Zeits. 70(3):275–299.
Stockmans F, Willi
ere Y. 1960. Hystrichosph
eres du
D
evonien belge (Sondage de l’Asile d’alien
es
a Tournai).
Sencken Lethaea 41:1–11.
Stockmans F, Willi
ere Y. 1962. Hystrichosph
eres du
D
evonien belge (Sondage de l’Asile d’alien
es
a Tournai).
Bull Soc Belge G
eol Pal
eontol Hydrol. 71:41–77.
Stockmans F, Willi
ere Y. 1963. Les Hystrichosph
eres ou
mieux les Acritarches du Silurien Belge. Sondage de la
Brasserie Lust
a Courtrai (Kortrijk). Bull Soc Belge G
eol
Pal
eontol Hydrol. 71(3):450–481.
Stolle E. 2007. Regional Permian palynological correlations:
Southeast Turkey-Northern Iraq. Comun Geol. 94:125–
143.
Stolle E. 2010. Recognition of southern Gondwanan palyno-
morphs at Gondwana’s northern margin and biostrati-
graphic correlation of Permian strata from SE Turkey
and Australia. Geol J. 45(2–3):336–349.
Stricanne L, Munnecke A, Pross J. 2006. Assessing mecha-
nisms of environmental change: Palynological signals
across the Late Ludlow (Silurian) positive isotope excur-
sion (d
13
C, d
18
O) on Gotland, Sweden. Palaeogeograph
Palaeoclimatol Palaeoecol. 230(1–2):1–31.
Strother PK. 1996. Acritarchs. In: Jansonius J, McGregor
DC, editors. Palynology: Principles and Applications.
Dallas (TX): American Association of Stratigraphic
Palynologists Foundation; p. 81–106.
Utting J. 1978. Geological Survey of Canada, Open File 593.
Ottawa: Natural Resources Canada, Geological Society
of Canada; p. 17.
Utting J, Spina A, Jansonius J, McGregor DC, Marshall
JEA. 2004. Reworked miospores in the Upper Paleozoic
and Lower Triassic of the northern circum-polar area
and selected localities. Palynology 28:75.
Utting J, Zonneveld JP, MacNaughton RB, Fallas KM,
Deunff J. 2005. Palynostratigraphy, lithostratigraphy
and thermal maturity of the Lower Triassic Toad
and Grayling, and Montney formations of western
Canada, and comparisons with coeval rocks of
the Sverdrup Basin, Nunavut. Bull Can Petrol Geol.
53(1):5–24.
Valensi L. 1953. Microfossiles des silex du Jurassique moyen:
Remarques p
etrographiques. M
em Soc G
eol France 32
(4):1–100.
Van de Schootbrugge B, Bailey TR, Rosenthal Y, Katz ME,
Wright JD, Miller KG, Feist-Burkhardt S, Falkowski
Palynology 343
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013
PG. 2005. Early Jurassic climate change and the radia-
tion of organic-walled phytoplankton in the Tethys
Ocean. Paleobiology 31(1):73–97.
Van de Schootbrugge B, Tremolada F, Rosenthal Y, Bailey
TR, Feist-Burkhardt S, Brinkhuis H, Pross J, Kent
DV, Falkowski PG. 2007. End-Triassic calcification
crisis and blooms of organic-walled ‘disaster species’.
Palaeogeograph Palaeoclimatol Palaeoecol. 244(1–4):
126–141.
Vavrdov
a M. 1966. Palaeozoic microplankton from central
Bohemia. Casopis Mineral Geol. 11(4):409–414.
Vavrdov
a M. 2008. Proterozoic acritarchs from the Precam-
brian-Cambrian transition in southern Moravia (M
ın
ın-1
borehole, Czech Republic). Bull Geosci. 83(1):85–92.
Visscher H, Brinkhuis H, Dilcher DL, Elsik WC, Eshet Y,
Looy CV, Rampino MR, Traverse A. 1996. The terminal
Paleozoic fungal event: evidence of terrestrial ecosystem
destabilization and collapse. Proceedings of the National
Academy of Sciences of the United States of America
93(5):2155–2158.
Visscher H, Brugman WA. 1981. Ranges of selected palyno-
morphs in the Alpine Triassic of Europe. Rev Palaeobot
Palynol. 34(1):115–128.
Visscher H, Sephton MA, Looy CV. 2011. Fungal virulence
at the time of the end-Permian biosphere crisis? Geology
39(9):883–886.
Vorob’eva NG, Sergeev VN, Knoll AH. 2009. Neoprotero-
zoic microfossils from the northeastern margin of the
East European Platform. J Paleontol. 83(2):161–196.
Wall D. 1965. Microplankton, pollen, and spores from
the Lower Jurassic in Britain. Micropaleontology
11(2):151–190.
Wall D, Downie C. 1963. Permian hystrichospheres from
Britain. Palaeontology 5(4):770–784.
Wang GQ, Xia WC. 2004. Conodont zonation across the
Permian Triassic boundary at the Xiakou section,
Yichang City, Hubei Province and its correlation with
the Global Stratotype Section and Point of the PTB. Can
J Earth Sci. 41(3):323–330.
Wilson LR. 1962. A Permian fungus spore type from the
Flowerpot Formation of Oklahoma. Okla Geol Notes
22(4):91–96.
Wood GD, Elsik WC. 1999. Paleoecologic and stratigraphic
importance of the fungus Reduviasporonites stoschianus
from the ‘Early-Middle’ Pennsylvanian of the Copaca-
bana Formation, Peru. Palynology 23:43–53.
Zonneveld JP, Beatty TW, MacNaughton RB, Pemberton
SG, Utting J, Henderson CM. 2010. Sedimentology and
ichnology of the Lower Triassic Montney Formation in
the Pedigree-Ring/Border-Kahntah River area, north-
western Alberta and northeastern British Columbia. Bull
Can Petrol Geol. 58(2):115–140.
344 Y. Lei et al.
Downloaded by [British Geological Survey], [Mr James B. Riding] at 07:20 15 November 2013