Fig 15 - uploaded by Tsuyoshi Hosoya
Content may be subject to copyright.
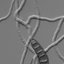
Urceolellapseudopani(TRL-818). A. Hairs with uncinate apex. That at upper right showing a narrow lumen nearly reaching the apex. Glassy portion indicated by gray tones. B. Asci. C. Section of apothecium showing ectal excipulum and hairs. D. Paraphyses of various types. Apex of that at left remained obtuse and not solidified; apices of three at right became tapered and solidified. E. Ascospores. Guttules shown only in the lower row. F. Schematic drawing of the apothecial section showing the outline of structure. Hairs not drawn. Scales. A-E, lOpm; F, 50~m.
Source publication







Fungal specimens of discomycetes in the family Hyaloscyphaceae were collected from across Japan and identified. In this report,
glassy members of the tribe Hyaloscypheae, subfamily Hyaloscyphoideae are described and illustrated. The 7 species involved
include two new species (Urceolella brunneola andU. pseudopani), and five species new to Japan (Hy...
Similar publications





Three Hyphodiscus species are described and illustrated: Hyphodiscus otanii sp. nov., Hyphodiscus hymeniophilus, and H. theiodeus, which is new to Japan. Culture studies revealed Phialophora-like anamorphs. Catenulifera gen. nov. is proposed for the anamorph of Hyphodiscus. The history of the genus is reviewed. Hyphodiscus can be delimited to membe...
Citations
... Gillet, Rodwayella Spooner, Sageria A. Funk and Tapesina Lambotte (Nag Raj and Kendrick 1975;Wang and Sutton 1984;Samuels and Rogerson 1990;Seifert et al. 2011;Crous et al. 2016aCrous et al. , b, 2018aGuatimosim et al. 2016). Connections of these anamorphs with their teleomorphs in Leotiomycetes were established only for a few species and limited to four genera such as Ascoconidium, Bloxamia, Chalara and Chaetochalara (Funk 1966a(Funk , b, 1975Nag Raj and Kendrick 1975;Hennebert and Bellemere 1979;Graddon 1980;Samuels and Rogerson 1990;Gams and Philippi 1992;Baral 1989Baral , 2002Hosoya and Otanl 1997;Paulin and Harrington 2000;Paulin et al. 2002;Guatimosim et al. 2016;Hosoya and Zhao 2016). The two known species of Ascoconidium, A. purpurascens and A. tsugae, were connected with Sageria in Helotiaceae (Funk 1966a(Funk , b, 1975. ...


The chalara-like anamorphs of Leotiomycetes are phialidic hyphomycetes with cylindrical collarettes and deeply seated sporulating loci, and hyaline, aseptate or septate, cylindrical conidia. They are commonly found on plant litters in both terrestrial and submerged environments, and with broad geographical distribution. This paper reports our research result of diversity, taxonomy and phylogeny of these fungi in China, which is based on a systematic study by using an integrated approach of literature study, morphological observation and phylogenetic analyses of 153 chalara-like fungal species with diversified morphology in conidiomata, setae, conidiophores, phialides and conidia. The phylogenetic analyses employing different datasets of SSU, LSU and ITS sequences of 116 species showed that these chalara-like fungi were paraphyletic and scattered in 20 accepted genera belonging to five families of Leotiomycetes: Arachnopezizaceae, Hamatocanthoscyphaceae, Helotiaceae, Neolauriomycetaceae and Pezizellaceae. Additional six genera, Ascoconidium , Bioscypha , Chalarodendron , Didonia , Phaeoscypha and Tapesina , all reported with chalara-like anamorphs in literatures, are also accepted as members of Pezizellaceae or Leotiomycetes genera incertae sedis. Among of these 26 accepted genera of chalara-like fungi in Leotiomycetes, 17 genera are asexually typified genera ( Ascoconidium , Bloxamia , Chalara , Chalarodendron , Constrictochalara , Cylindrochalara , Cylindrocephalum , Leochalara , Lareunionomyces , Minichalara , Neochalara , Neolauriomyces , Nagrajchalara , Parachalara , Stipitochalara , Xenochalara and Zymochalara ), and 9 are sexually typified genera ( Bioscypha , Bloxamiella , Calycellina , Calycina , Didonia , Hymenoscyphus , Mollisina , Phaeoscypha and Tapesina ). The phylogenetic significance of conidial septation in generic delimitation was further confirmed; while other morphologies such as conidiomata, setae, conidiophores, phialides, conidial length, and conidial ornamentation have little phylogenetic significance, but could be used for species delimitation. The polyphyletic genus Chalara s. lat. is revised with monophyletic generic concepts by redelimitation of Chalara s. str. in a narrow concept, adaption of the emended Calycina to also include asexually typified chalara-like fungi, reinstatement of Cylindrocephalum , and introduction of six new genera: Constrictochalara W.P. Wu & Y.Z. Diao, Leochalara W.P. Wu & Y.Z. Diao, Minichalara W.P. Wu & Y.Z. Diao, Nagrajchalara W.P. Wu & Y.Z. Diao, Parachalara W.P. Wu & Y.Z. Diao and Stipitochalara W.P. Wu & Y.Z. Diao. Chaetochalara becomes a synonym of Chalara s. str., and the known species are disassembled into Chalara s. str. and Nagrajchalara . The polyphyletic genus Bloxamia is also redefined by introducing the new genus Bloxamiella W.P. Wu & Y.Z. Diao for B . cyatheicola . Five existing species of Chalara s. lat. were excluded from Leotiomycetes and reclassified: Chalara breviclavata as Chalarosphaeria breviclavata W.P. Wu & Y.Z. Diao gen. et sp. nov. in Chaetosphaeriaceae, C . vaccinii as Sordariochalara vaccinii W.P. Wu & Y.Z. Diao gen. et sp. nov. in Lasiosphaeriaceae, and three other Chalara species with hyaline phialides, C . hyalina , C . schoenoplecti and C . siamense as combinations of Pyxidiophora in Pyxidiophoraceae. For biodiversity of these fungi in China, a total of 80 species in 12 genera, including 60 new species, 17 new records and 1 new name, were discovered and documented in this paper. In addition, five species including three new species are reported from Japan. In connection to this revision, a total of 44 new combinations are made. The identification keys are provided for most of these genera. Future research area of these fungi should be the phylogenetic relationship of several sexually typified genera such as Bioscypha , Calycellina , Calycina , Didonia , Phaeoscypha , Rodwayella and Tapesina , and systematic revision of existing names under the genera Bloxamia , Chaetochalara and Chalara .
... The morphology of Hyphodiscus cajaniensis agrees with the concept of Hyphodiscus used here as well as in previous literature. Notes on Hyphopeziza: Selected publications chronologically arranged are Svrček (1985), Huhtinen (1987), Hosoya & Otani (1997b), and Han et al. (2014). This monotypic genus was erected by Han et al. (2014) during their revision of Hyaloscyphaceae. ...
... The name Hyphopeziza was created in reference to its resemblance to both Hyalopeziza and Hyphodiscus. It grows on angiosperm leaves (Quercus, Carpinus) in Europe and Asia (Japan, Korea) (Table 2, Huhtinen 1987, Hosoya & Otani 1997b, Han et al. 2014). According to Han et al. (2014) the genus is characterised by white-greyish, minutely pubescent apothecia, 0.5-1 mm in diam The ectal excipulum is hyaline to pale brown textura (globulosa-) angularis to t. prismatica, slightly gelatinised, cells thin-to slightly thick-walled. ...
... It was isolated from acidic soils in Czech Republic (Hujslová et al. 2014). Hosoya & Otani (1997b) briefly described cultures of Hyphopeziza, but they did not find yeast-like colonies described by Hujslová et al. (2014). Given that the cultures of Soosiella and Hyphopeziza are both sterile they are difficult to compare. ...





In a recently published classification scheme for Leotiomycetes, the new family Hyphodiscaceae was erected; unfortunately, this study was rife with phylogenetic misinterpretations and hampered by a poor understanding of this group of fungi. This manifested in the form of an undiagnostic familial description, an erroneous familial circumscription, and the redescription of the type species of an included genus as a new species in a different genus. The present work corrects these errors by incorporating new molecular data from this group into phylogenetic analyses and examining the morphological features of the included taxa. An emended description of Hyphodiscaceae is provided, notes and descriptions of the included genera are supplied, and keys to genera and species in Hyphodiscaceae are supplied. Microscypha cajaniensis is combined in Hyphodiscus, and Scolecolachnum nigricans is a taxonomic synonym of Fuscolachnum pteridis. Future work in this family should focus on increasing phylogenetic sampling outside of Eurasia and better characterising described species to help resolve outstanding issues.
Citation: Quijada L, Baral HO, Johnston PR, Pärtel K, Mitchell JK, Hosoya T, Madrid H, Kosonen T, Helleman S, Rubio E, Stöckli E, Huhtinen S, Pfister DH (2022). A review of Hyphodiscaceae. Studies in Mycology 103: 59–85. doi: 10.3114/sim.2022.103.03
... However, we expected more undocumented members. In a series of studies, we contributed to the enumeration of undocumented taxa, including new species (Hosoya & Otani, 1997a, 1997bHosoya & Harada, 1999;Tochihara & Hosoya, 2019). Cantrell and Hanlin (1997) first conducted a phylogenetic analysis to assess the taxonomy of Hyaloscyphaceae using a single gene, ITS-5.8S. ...
... Otani (1989) enumerated the Japanese mycobiota of Discomycetes, a taxon no longer used but still practical in the modern taxonomy, and enumerated 77 genera with 214 species in the order. Through our exploration and enumeration, we added 57 newly documented taxa in Japan, and 30 new taxa, including 2 genera (Otani, Hosoya, & Furuya, 1991;Hosoya et al, 1993;Kubono & Hosoya, 1994;Hosoya & Otani 1995, 1997a, 1997b, 1997cHosoya & Harada 1999;Hosoya, 2000Hosoya, , 2002Hosoya, , 2004Hosoya, , 2005Hosoya, , 2009Hosoya & Huhtinen 2002;Narumi-Saito, Hosoya, Sano, & Harada, 2006;Hosoya et al., 2010aHosoya et al., , 2010cHosoya et al., , 2011Hosoya, Sung, Han, & Shin, 2010d;Hosoya et al., 2012;Hosoya, Hosaka, Saito, Degawa, & Suzuki, 2013a;Hosoya et al., 2013b;Hosoya, Saito, & Sasagawa, 2013c;Zhao, Hosoya, Shirouzu, Kakishima, & Yamaoka, 2013;Han et al., 2014a;Hosoya, Zhao, & Degawa, 2014;Degawa et al., 2015;Gross, Hosoya, Zhao, & Baral, 2015;Itagaki, Nakamura, & Hosoya, 2019;Tochihara & Hosoya, 2019). These included new species, new genera of both sexual and asexual states, now being merged under single names (some examples are presented in Fig. 2). ...

Helotiales is one of the most diverse groups of apothecial ascomycetes, including 3000—4000 taxa. Recent progress in the systematics, ecology, and their applications through research is herein reviewed based on the experiences of the author with a special emphasis on activities in Japan. In the past 30 y, more than 50 helotialean taxa have been added to the mycobiota of Japan, including new taxa. With the advent of molecular phylogeny, some families have been revisited, such as members with stroma (Sclerotiniaceae and Rutstroemiaceae) or hairs (Hyaloscyphaceae and Lachnaceae). Although the monophyly of Helotiales has not yet been demonstrated, our understanding of its phylogeny has greatly advanced. The unexpected ecological nature represented by endophytism has been revealed through barcoding and other molecular techniques. The research history of ash dieback is also reviewed, and the endophytism/saprophytism of the pathogen on its original host is discussed. Drug discoveries within Helotiales are reviewed, and successful examples are presented. As future perspectives, both the cumulation of occurrence and sequence data of Helotiales is greatly encouraged to elucidate this important group of fungi.
... HC4 root endophyte sp. 1e3 secreted droplets on the colony, as observed in related Hyaloscyphaceae fungal species (Hosoya & Harada, 1999;Hosoya & Otani, 1997). The dispersal mechanism of some helotialean root endophytes has long been a mystery for researchers (Brenn, Menkis, Grünig, Sieber, & Holdenrieder, 2008;Queloz et al., 2005;Stroheker, Queloz, & Sieber, 2016). ...
We used a culture-dependent approach to investigate root-associated fungal communities in Fagaceae roots at four fagaceous species-dominant forests in Japan. In total 1029 isolates were collected and classified, based on colony morphological features and molecular information. Species of order Helotiales (Ascomycota) were dominantly isolated at all four sites, in which a globally-distributed putative endophytic group in Hyaloscyphaceae predominated. This group of fungi was morphologically and phylogenetically investigated using these isolates as well as additional isolates collected from 8 different sites in Japan. Among the Hyaloscyphaceae, Glutinomyces species were frequently detected, and three novel species, G. inflatus, G. vulgaris, and G. takaragaikensis spp. nov. were identified and described according to their morphology and genealogical concordance.
... As shown in the phylogenies (Figs 2 and 3), H. pygmaea was not grouped with Hyalopeziza spp. (Clade 5), suggesting that this taxon has been misclassified (Huhtinen 1987;Hosoya & Otani 1997). Although H. pygmaea was nested within Hyphodiscus spp. ...


Hyaloscyphaceae is the largest family in Helotiales, Leotiomycetes. It is mainly characterized by minute apothecia with well-differentiated hairs, but its taxonomic delimitation and infrafamilial classification remain ambiguous. This study performed molecular phylogenetic analyses using multiple genes including the ITS-5.8S rDNA, the D1-D2 region of large subunit of rDNA, RNA polymerase II subunit 2, and the mitochondrial small subunit. The primary objective was to evaluate the phylogenetic utility of morphological characters traditionally used in the taxonomy of Hyaloscyphaceae through reassessment of the monophyly of this family and its genera. The phylogenetic analyses inferred Hyaloscyphaceae as being a heterogeneous assemblage of a diverse group of fungi and not supported as monophyletic. Among the three tribes of Hyaloscyphaceae only Lachneae formed a monophyletic lineage. The presence of hairs is rejected as a synapomorphy, since morphologically diversified hairs have originated independently during the evolution of Helotiales. The true- and false-subiculum in Arachnopezizeae are hypothesized to have evolved through different evolutionary processes; the true-subiculum is likely the product of a single evolutionary origin, while the false-subiculum is hypothesized to have originated multiple times. Since Hyaloscyphaceae sensu lato was not resolved as monophyletic, Hyaloscyphaceae sensu stricto is redefined and only applied to the genus Hyaloscypha.
... Otani (1989) published a list of Japanese discomycetes which included 41 taxa of Hyaloscyphaceae to prepare a future volume of discomycetes in " Fungal flora of Japan " . Since then, based on collections throughout Japan (Fig. 2), 29 taxa including 10 newly described species have been added to the list (Hosoya, 2002Hosoya, , 2004 Hosoya & Harada, 1999; Hosoya & Huhtinen, 2002; Hosoya & Otani, 1997a, 1997b; Ono and Hosoya, 2001; Tanaka and Hosoya, 2001). As has been stated by Korf (1958), " for the discosystematist, Japan is a relatively unexplored paradise " . ...
Due to the regional broadening in south and north, Japan embraces a wide range of diver-sified organisms, and Fungi are not the exception. In spite of relatively long historical background in inventory of Japan's mycobiota, first compiled in 1905, much information remains to be added. Cur-rently, rearrangement and databasing of the specimens in the mycological herbarium in National Sci-ence Museum (TNS) have been carried out, and some of the data is publically available. To extend the collection, strategic collection building, based on analysis of the specimens at hand and collabora-tion with other parties, are required. GBIF, an international biological databasing project, provides a good opportunity to integrate information of specimens conserved in the major mycological herbaria in Japan.
... Monoconidial isolation was done on agar-leaf piece media using the diseased leaves of Japanese barnyard millet (Furukawa and Kishi 2002;Sato and Moriwaki 2003). A germinated conidium was transferred onto modifi ed Weitzman and Silva-Hunter (mWSH) agar (Weitzman and Silva-Hunter 1967;Hosoya and Otani 1997) for each isolate. The isolates were maintained on mWSH slants and deposited in National Institute of Agrobiological Sciences (NIAS) Genebank (MAFF) in Japan as MAFF 511471 to 511473. ...
Five isolates of a species of Colletotrichum were collected from Japanese barnyard millet (Echinochloa utilis) in Japan. Although the fungus had once been identified as C. graminicola sensu lato, it was clearly different from C. graminicola isolated from maize (Zea mays) in its falcate and short conidia, 18.0–22.2μm in length, cultural characteristics, and specific pathogenicity to E. utilis. Moreover, molecular phylogenetic analyses using sequences of rDNA-ITS, HMG, and SOD2 indicated a monophyly of the isolates. A new species, Colletotrichum echinochloae, is then proposed based on the morphological, pathological, and molecular characteristics.
... Also found on the bases of the stems of some of the collections are apothecia of yet another inoperculate discomycete, Hyalopeziza millepunctata (Lib.) Raitv., a species recently critically revised by Hosoya & Otani (1997). The cultural studies reported by Yafiez-Morales et al. (1998) demonstrate that the sclerotial fungus is distinct from either of the discomycetes on the same stems. ...


A new species of Crocicreas is reported on overwintered stems of Epifagus virginianus in New York State in apparently non-organic association with sclerotia of Sclerotium orobanches, itself a rarely reported fungus. The changing concepts of the generic names Crocicreas and Cyathicula are discussed in regard to placement of the new taxon.

The circumscription and composition of the Hyaloscyphaceae are controversial and based on poorly sampled or unsupported phylogenies. The generic limits within the hyaloscyphoid fungi are also very poorly understood. To address this issue, a robust five-gene Bayesian phylogeny (LSU, RPB1 , RPB2 , TEF-1α , mtSSU; 5521 bp) with a focus on the core group of Hyaloscyphaceae and Arachnopezizaceae is presented here, with comparative morphological and histochemical characters. A wide representative sampling of Hyaloscypha supports it as monophyletic and shows H. aureliella (subgenus Eupezizella ) to be a strongly supported sister taxon. Reinforced by distinguishing morphological features, Eupezizella is here recognised as a separate genus, comprising E. aureliella , E. britannica , E. roseoguttata and E. nipponica (previously treated in Hyaloscypha ). In a sister group to the Hyaloscypha-Eupezizella clade a new genus, Mimicoscypha , is created for three seldom collected and poorly understood species, M. lacrimiformis , M. mimica ( nom. nov. ) and M. paludosa , previously treated in Phialina , Hyaloscypha and Eriopezia , respectively. The Arachnopezizaceae is polyphyletic, because Arachnoscypha forms a monophyletic group with Polydesmia pruinosa , distant to Arachnopeziza and Eriopezia ; in addition, Arachnopeziza variepilosa represents an early diverging lineage in Hyaloscyphaceae s.str. The hyphae originating from the base of the apothecia in Arachnoscypha are considered anchoring hyphae (vs a subiculum) and Arachnoscypha is excluded from Arachnopezizaceae . A new genus, Resinoscypha , is established to accommodate Arachnopeziza variepilosa and A. monoseptata , originally described in Protounguicularia . Mimicoscypha and Resinoscypha are distinguished among hyaloscyphoid fungi by long tapering multiseptate hairs that are not dextrinoid or glassy, in combination with ectal excipulum cells with deep amyloid nodules. Unique to Resinoscypha is cyanophilous resinous content in the hairs concentrated at the apex and septa. Small intensely amyloid nodules in the hairs are furthermore characteristic for Resinoscypha and Eupezizella . To elucidate species limits and diversity in Arachnopeziza , mainly from Northern Europe, we applied genealogical concordance phylogenetic species recognition (GCPSR) using analyses of individual datasets (ITS, LSU, RPB1 , RPB2 , TEF-1α ) and comparative morphology. Eight species were identified as highly supported and reciprocally monophyletic. Four of these are newly discovered species, with two formally described here, viz. A. estonica and A. ptilidiophila . In addition, Belonium sphagnisedum , which completely lacks prominent hairs, is here combined in Arachnopeziza , widening the concept of the genus. Numerous publicly available sequences named A. aurata represent A. delicatula and the confusion between these two species is clarified. An additional four singletons are considered to be distinct species, because they were genetically divergent from their sisters. A highly supported five-gene phylogeny of Arachnopezizaceae identified four major clades in Arachnopeziza , with Eriopezia as a sister group. Two of the clades include species with a strong connection to bryophytes; the third clade includes species growing on bulky woody substrates and with pigmented exudates on the hairs; and the fourth clade species with hyaline exudates growing on both bryophytes and hardwood. A morphological account is given of the composition of Hyaloscyphaceae and Arachnopezizaceae , including new observations on vital and histochemical characters.
This article has been withdrawn at the request of the author(s) and/or editor. The Publisher apologizes for any inconvenience this may cause.
The full Elsevier Policy on Article Withdrawal can be found at https://www.elsevier.com/about/our-business/policies/article-withdrawal