Fig 6 - uploaded by Dominik Begerow
Content may be subject to copyright.

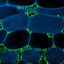
Ultrastructural characteristics of the Classiculomycetes (a, b) and Cryptomycocolacomycetes (c-f). Bars = 0.1 μm in a-c, f, 0.2 μm in d, and 1 μm in e a Septal pore apparatus of Classicula fluitans R. Bauer, Begerow, Oberw. & Marvanová ATCC 64713 with nonswollen pore margin and associated microbodies (arrowheads) in a more or less circular arrangement. Note that the organelle-free zone is of poor contrast b Intranuclear mitotic metaphase SPB of Jaculispora submersa H.J. Huds. & Ingold CCM 8127; nuclear envelope (arrowheads), mitochondria (m) c Simple septal pore of Cryptomycocolax abnormis Oberw. & R. Bauer FO 40023 asso-
Source publication








The simple-septate basidiomycetes comprise more than 8,000 species that show a high morphological and ecological heterogeneity. To gain insight in the phylogenetic relationships within this group, we compared several ultrastructural features such as septal pore apparatus, form, and behavior of the spindle pole bodies, types of host–parasite interac...
Contexts in source publication
Context 1
... Jülich Septal pores with microbodies ( Fig. 5a Fig. 6e,f), septal pores with microbodies ( Fig. 6c), metaphasic SPBs in the nuclear envelope (Fig. 6d) Cystobasidiaceae Gäum. With characters of the ...
Context 2
... Jülich Septal pores with microbodies ( Fig. 5a Fig. 6e,f), septal pores with microbodies ( Fig. 6c), metaphasic SPBs in the nuclear envelope (Fig. 6d) Cystobasidiaceae Gäum. With characters of the ...
Context 3
... Jülich Septal pores with microbodies ( Fig. 5a Fig. 6e,f), septal pores with microbodies ( Fig. 6c), metaphasic SPBs in the nuclear envelope (Fig. 6d) Cystobasidiaceae Gäum. With characters of the ...
Context 4
... septal pore apparatus in both species is composed of a simple pore surrounded by microbodies in a circular arrangement ( Fig. 6a; Bauer et al. 2003). In basidiomycetes, this type of septal pore apparatus occurs only in the Figs. 6c and 10a; Bauer and Oberwinkler 1994;Kirschner et al. 2001;Oberwinkler and Bauer 1990), it occurs sporadically also in some members of the Atractiellomycetes. Thus, Saccoblastia in the Atractiellomycetes possesses this septal pore type ...
Context 5
... R. Bauer and Colacosiphon filiformis R. Kirscher, R. Bauer & Oberw. form a well-supported clade ( Fig. 1 and Table 1). This phylogenetic placement agrees well with morphological, ultrastructural, and ecological results. Both fungi have uniquely long basidia, and they are both mycoparasites, interacting with their ascomycetous hosts by colacosomes (Fig. 6e,f; Kirschner et al. 2001;Oberwinkler and Bauer 1990). In addition, the septal pore apparatus in Cryptomycocolax is essentially identical to that of Colacosiphon: in both species the septal pores are surrounded by microbodies and bodies resem- bling Woronin bodies (Fig. 6c). Like in ascomycetes, the SPB discs in Cryptomycocolax ...
Context 6
... mycoparasites, interacting with their ascomycetous hosts by colacosomes (Fig. 6e,f; Kirschner et al. 2001;Oberwinkler and Bauer 1990). In addition, the septal pore apparatus in Cryptomycocolax is essentially identical to that of Colacosiphon: in both species the septal pores are surrounded by microbodies and bodies resem- bling Woronin bodies (Fig. 6c). Like in ascomycetes, the SPB discs in Cryptomycocolax (Colacosiphon has not been investigated in this respect) are large in size, they are inserted into an otherwise intact nuclear envelope (Fig. 6d), and they duplicate by fission ( Oberwinkler and Bauer 1990). Thus, SPB and septal pore features indicate a very basal phylogenetic ...
Context 7
... identical to that of Colacosiphon: in both species the septal pores are surrounded by microbodies and bodies resem- bling Woronin bodies (Fig. 6c). Like in ascomycetes, the SPB discs in Cryptomycocolax (Colacosiphon has not been investigated in this respect) are large in size, they are inserted into an otherwise intact nuclear envelope (Fig. 6d), and they duplicate by fission ( Oberwinkler and Bauer 1990). Thus, SPB and septal pore features indicate a very basal phylogenetic position of the Cryptomycocolacomy- cetes within the ...
Context 8
... that in the molecular analyses the representatives of the Cryptomycocolacomycetes are separated from the Microbotryomycetes (Fig. 1). How- ever, a closer inspection reveals high ultrastructural divergence between the Cryptomycocolacomycetes and Microbotryomycetes. In fact, the puccinialean septal pores of Cryptomycocolax and Colacosiphon (Fig. 6c) are similar to the septal pores of Saccoblastia of the Atractiellomycetes (Fig. 5a), to those of most Puccinio- mycetes ( Fig. 10a; see Bauer and Oberwinkler 1994 and the references therein) and the members of the Classiculomycetes ( Fig. 6a; Bauer et al. 2003), but definitely not to those of the Microbotryomycetes (Fig. 8a-c; ...
Context 9
... 2003), but definitely not to those of the Microbotryomycetes (Fig. 8a-c; Oberwinkler and Bauer 1990;Kirschner et al. 1999;Oberwinkler et al. 1990a,b). Furthermore, Cryptomycocolax (Colacosiphon has not been investi- gated in this respect) the discoidal SPBs are inserted in a closely fitting pore of the nuclear envelope during nuclear division ( Fig. 6d; Oberwinkler and Bauer 1990). Cryptomycocolax shares this feature with the Puccinio- mycetes ( Fig. 10b; Bauer 1987;Oberwinkler 1990a, 1994;Boehm and McLaughlin 1989;Bourett and McLaughlin 1986;O'Donnell and McLaughlin 1981a-c) and with the Atractiellomycetes (Fig. 5c), but not with the Microbotryomycetes ( Fig. 8d; Berbee et al. ...
Context 10
... Cryptomycocolax (Colacosiphon has been unsuffi- ciently investigated in this respect) a second type of colacosome was found along the cytoplasmic intrusions of the host formed into the hyphae of the parasite (Fig. 6e,f; Oberwinkler and Bauer 1990). Colacosomes of that type have a more electron-transparent core, and they are capable of fusing with host cells via a small pore. Fusion with the host cytoplasm may be the ancestral function of colaco- somes, distinguishing the mycoparasitic behavior of Cryptomycocolax (Cryptomycocolacomycetes) from that of ...
Context 11
... 1990). Colacosomes of that type have a more electron-transparent core, and they are capable of fusing with host cells via a small pore. Fusion with the host cytoplasm may be the ancestral function of colaco- somes, distinguishing the mycoparasitic behavior of Cryptomycocolax (Cryptomycocolacomycetes) from that of the Microbotryomycetes (compare Fig. 6e,f with Fig. 8e). Accordingly, the evolution of the Microbotryo- mycetes may have been accompanied by the loss of the colacosome-fusion interaction. In addition, the question arises whether or not the colacosome-fusion mycoparasit- ism of Cryptomycocolax is homologous to the nanometer- fusion interaction occurring in the Cystobasidiomycetes, ...
Context 12
... may have been accompanied by the loss of the colacosome-fusion interaction. In addition, the question arises whether or not the colacosome-fusion mycoparasit- ism of Cryptomycocolax is homologous to the nanometer- fusion interaction occurring in the Cystobasidiomycetes, Agaricostilbomycetes and possibly also in the Classiculo- mycetes (compare Fig. 6f with Fig. 4c-f and Fig. 7d-f; see also Bauer 2004 and the discussion of the Agaricostilbo- mycetes and Cystobasidiomycetes). It is plausible that the colacosome-fusion interaction occurring in Cryptomycoco- lax represents the ancestral type of the nanometer-fusion interaction. In a first evolutionary step, an apoplastic compartment, ...
Context 13
... In view of the known fixation problems with these species, these structures might in fact be micro- bodies that artificially lack the surrounding membrane. Outside the Pucciniomycetes, septal pores associated with microbodies occur also in the Saccoblastiaceae of the Atractiellomycetes ( Fig. 5a; Weiß et al. 2004), in the Cryptomycocolacales ( Fig. 6c; Oberwinkler and Bauer 1990) and in the Classiculomycetes ( Fig. 6a; Bauer et al. ...
Context 14
... structures might in fact be micro- bodies that artificially lack the surrounding membrane. Outside the Pucciniomycetes, septal pores associated with microbodies occur also in the Saccoblastiaceae of the Atractiellomycetes ( Fig. 5a; Weiß et al. 2004), in the Cryptomycocolacales ( Fig. 6c; Oberwinkler and Bauer 1990) and in the Classiculomycetes ( Fig. 6a; Bauer et al. ...
Context 15
... examined, the discoidal SPBs are inserted in a close-fitting pore of the nuclear envelope during nuclear division ( Fig. 10b; Bauer 1987;Bauer and Oberwinkler 1994;Boehm and McLaughlin 1989;Bourett and McLaughlin 1986;O'Donnell and McLaughlin 1981a-c). This SPB-nuclear envelope relationship is shared only with the Crytomyco- colacales ( Fig. 6d; Oberwinkler and Bauer 1990) and the Atractiellales ( Fig. 5c; McLaughlin 1987). In Pachnocybe ( Oberwinkler 1990a), Herpobasidium (Bauer and, Helicobasidium (Bourett andMcLaughlin 1986) andEocronartium (Boehm and, but definitely not in the rusts ( Fig. 10b; Bauer 1987;O'Donnell and McLaughlin 1981a-c), a characteristic cap of ...
Citations
... For several groups a typical mycoparasite is illustrated. The relationships depicted in this phylogram represent an interpretation based on multiple previously published phylogenetic reconstructions (Aime et al. 2006, Bauer et al. 2006, Wang et al. 2015a 4. Discussion of groups in systematic order ...
... In literature, Agaricostilbomycetes are often regarded as lacking haustorial cells (Bauer et al. 2006). However, Seifert et al. (1992) reported the development of haustorial cells in Stilbum vulgare attaching to hyphae of Cladosporium sp. when growing in co-culture. ...
... These outgrows may reach up to 20 μm in length and bifurcation has been observed occasionally. Bauer et al. (2006) reported the attachment of these filamentous outgrows to hyphae of the own mycelium, and interpreted this as self-parasitism, possibly being a mechanism to redistribute nutrients in the own mycelium. ...


Among the over 31,000 recognized species within Basidiomycota, approximately 200 are documented as parasites of other fungi, so-called mycoparasites. They exhibit a remarkable diversity in morphological characteristics, host-parasite interaction structures, and phylogenetic affiliations. Mycoparasitism as a trophic strategy in Basidiomycota has received little attention from the mycological community, with most research performed during the last decades of the previous century. These mycoparasites are documented across three subphyla, encompassing 10 different classes, yet a comprehensive understanding of their evolutionary relationships and host-parasite interaction mechanisms remains limited, and much data are scattered in the literature. This chapter aims to systematically outline the various classes of Basidiomycota known to include mycoparasites, elucidating their diversity, host-parasite interface, life cycle, morphology, and systematic classification. To provide a comprehensive overview of current knowledge on basidiomycetous mycoparasitism, we have compiled a catalogue of presumed mycoparasites, detailing available data for each species.
... Molecular phylogenies have revealed that mycoparasitism mainly occurs in two subphyla of Basidiomycota: Agaricomycotina and Pucciniomycotina (Fig. 1) (Weiß et al. 2004, Bauer et al. 2006, Begerow et al. 2017. In Agaricomycotina, the majority of mycoparasites are members of Tremellomycetes, whereas only few belong to Agaricomycetes, e.g., species of Asterophora, Pseudoboletus, and Squamanita (Redhead et al. 1994, Oberwinkler 2012, Weiß et al. 2014, Koch & Herr 2021, Caiafa & Smith 2022. ...
... The occurrence of mycoparasitism in Tritirachiomycetes (Pucciniomycotina) was suggested by Aime et al. (2014), although no cellular interaction structures or specific mechanisms for nutrient transfer were reported (Beguin 2010). (Aime et al. 2006, Bauer et al. 2006, Schell et al. 2011, Wang et al. 2015a, Zhao et al. 2017, He et al. 2019. Names of classes indicated in red represent those comprising mycoparasitic species. ...
... Two major interaction mechanisms have been described: (i) fusion-interaction and (ii) colacosome-interaction (Oberwinkler & Bauer 2018). A large variation at the ultrastructural level exists within each of these interaction types (Bauer 2004, Bauer et al. 2006, Oberwinkler & Bauer 2018). ...

Mycoparasites in Basidiomycota comprise a diverse group of fungi, both morphologically and phylogenetically. They interact with their hosts through either fusion-interaction or colacosome-interaction. Colacosomes are subcellular structures formed by the mycoparasite at the host–parasite interface, which penetrate the parasite and host cell walls. Previously, these structures were detected in 19 fungal species, usually by means of transmission electron microscopy. Most colacosome-forming species have been assigned to Microbotryomycetes ( Pucciniomycotina , Basidiomycota ), a highly diverse class, comprising saprobic yeasts, mycoparasites, and phytoparasites. In general, these myco- and phytoparasites are dimorphic organisms, with a parasitic filamentous morph and saprobic yeast morph. We investigated colacosome-forming mycoparasites based on fungarium material, freshly collected specimens, and cultures of yeast morphs. We characterised the micromorphology of filamentous morphs, the physiological characteristics of yeast morphs, and inferred phylogenetic relationships based on DNA sequence data from seven loci. We outline and employ an epifluorescence-based microscopic method to assess the presence and organisation of colacosomes. We describe five new species in the genus Colacogloea , the novel dimorphic mycoparasite Mycogloiocolax gerardii , and provide the first report of a sexual, mycoparasitic morph in Colacogloea philyla and in the genus Slooffia . We detected colacosomes in eight fungal species, which brings the total number of known colacosome-forming fungi to 27. Finally, we revealed three distinct types of colacosome organisation in Microbotryomycetes .
... Molecular phylogenies have revealed that mycoparasitism mainly occurs in two subphyla of Basidiomycota: Agaricomycotina and Pucciniomycotina (Fig. 1) (Weiß et al. 2004, Bauer et al. 2006, Begerow et al. 2017. In Agaricomycotina, the majority of mycoparasites are members of Tremellomycetes, whereas only few belong to Agaricomycetes, e.g., species of Asterophora, Pseudoboletus, and Squamanita (Redhead et al. 1994, Oberwinkler 2012, Weiß et al. 2014, Koch & Herr 2021, Caiafa & Smith 2022. ...
... The occurrence of mycoparasitism in Tritirachiomycetes (Pucciniomycotina) was suggested by Aime et al. (2014), although no cellular interaction structures or specific mechanisms for nutrient transfer were reported (Beguin 2010). (Aime et al. 2006, Bauer et al. 2006, Schell et al. 2011, Wang et al. 2015a, Zhao et al. 2017, He et al. 2019. Names of classes indicated in red represent those comprising mycoparasitic species. ...
... Two major interaction mechanisms have been described: (i) fusion-interaction and (ii) colacosome-interaction (Oberwinkler & Bauer 2018). A large variation at the ultrastructural level exists within each of these interaction types (Bauer 2004, Bauer et al. 2006, Oberwinkler & Bauer 2018). ...
... This situation has shaped the evolution of these organisms through host specialization and, most likely, genome expansion (2). These traits are relevant for distinguishing the Pucciniales from their sister taxa, along with the absence of a cap of endoplasmic reticulum enclosing the spindle pole body discs during nuclear division (3). Furthermore, the Pucciniales, along with their sister orders (i.e., the class Pucciniomycetes), are notorious for undergoing mitosis and cell division without the formation of clamp connections (in most Basidiomycota, clamp connections ensure the maintenance of the heterokaryon upon cell division). ...
... Furthermore, the Pucciniales, along with their sister orders (i.e., the class Pucciniomycetes), are notorious for undergoing mitosis and cell division without the formation of clamp connections (in most Basidiomycota, clamp connections ensure the maintenance of the heterokaryon upon cell division). Finally, Pucciniomycotina can be distinguished from the other subphyla in the Basidiomycota by cell wall sugar composition and traits in the septal pores and by the presence of disc-like spindle pole bodies (3,4). ...



Within Eukaryotes, fungi are the typical representatives of haplontic life cycles. Basidiomycota fungi are dikaryotic in extensive parts of their life cycle, but diploid nuclei are known to form only in basidia. Among Basidiomycota, the Pucciniales are notorious for presenting the most complex life cycles, with high host specialization, and for their expanded genomes. Using cytogenomic (flow cytometry and cell sorting on propidium iodide-stained nuclei) and cytogenetic (FISH with rDNA probe) approaches, we report the widespread occurrence of replicating haploid and diploid nuclei (i.e., 1C, 2C and a small proportion of 4C nuclei) in diverse life cycle stages (pycnial, aecial, uredinial, and telial) of all 35 Pucciniales species analyzed, but not in sister taxa. These results suggest that the Pucciniales life cycle is distinct from any cycle known, i.e., neither haplontic, diplontic nor haplodiplontic, corroborating patchy and disregarded previous evidence. However, the biological basis and significance of this phenomenon remain undisclosed.
IMPORTANCE Within Eukaryotes, fungi are the typical representatives of haplontic life cycles, contrasting with plants and animals. As such, fungi thus contain haploid nuclei throughout their life cycles, with sexual reproduction generating a single diploid cell upon karyogamy that immediately undergoes meiosis, thus resuming the haploid cycle. In this work, using cytogenetic and cytogenomic tools, we demonstrate that a vast group of fungi presents diploid nuclei throughout their life cycles, along with haploid nuclei, and that both types of nuclei replicate. Moreover, haploid nuclei are absent from urediniospores. The phenomenon appears to be transversal to the organisms in the order Pucciniales (rust fungi) and it does not occur in neighboring taxa, but a biological explanation or function for it remains elusive.
... Agarics are mushroom-forming fungi also called euagarics and their hymenium is formed on gills. They belong to the subdivision Agaricomycotina, class Agaricomycetes (Moncalvo et al., 2002;Bauer et al., 2006). They produce important natural substances used in agriculture (e.g., strobilurines), medicine (e.g., pleuromutilines), and biotechnology (e.g., polysaccharides; Pointing et al., 2001;Webster and Weber, 2007;Kück et al., 2014;Hyde et al., 2019;Sandargo et al., 2019). ...




... Hence, molecular techniques are recognized to be a powerful aid for traditional taxonomic methods in identifying mushrooms. The main application was in food quality control and the pharmaceutical industry (Bauer et al. 2006). ...


Humans use mushrooms for food, medicine, poison, and other purposes. Traditionally, mushrooms are used to treat several diseases locally. Treatment knowledge using mushrooms exists in the local communities. The wealth of ethnomedicinal knowledge is directly proportional to the diversity of mushroom species in a said geographical area. Western Ghats of India, one of the global biodiversity hot spots, is the home to diverse ethnomedicinal mushrooms. Mushroom diversity is responsible for the valuable ethnomedicinal knowledge throughout the Western Ghats. Locally, mushrooms are collected from the wild for food and medicine. Ethnomedicinal mushrooms are gaining importance because of their established pharmacological evidence as medicines, nutraceuticals, and food supplements. Even now, most of the mushrooms are collected from the wild because only few of them have been commercialized using modern techniques. The unsustainable collection, habitat degradation because of human interference, and climate change pose a great threat to wild ethnomedicinal mushrooms. Hence, conservation and commercialization of potent ethnomedicinal mushrooms are the need of the hour.Keywords
Basidiomycota
EthnomedicineHuman healthMushroomsThe Western Ghats
... Одним із результатів проведених досліджень стало виокремлення монотипного роду Naohidea Oberw. (Oberwinkler, 1990), самостійність якого згодом була підтверджена молекулярногенетичними методами (Sampaio et al., 1999;Bauer et al., 2006). Рід був названий на честь японського урединіолога, професора Наохіде Хіратсуки (Oberwinkler, 1990). ...
... із класу Cystobasidiomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. (Bauer et al., 2006;Denchev, 2009). Геном N. sebacea було секвеновано лише нещодавно (JGI, 2022). ...
... Naohidea sebacea є диморфним грибом, у життєвому циклі якого представлені гаплоїдна дріжджова та дикаріотична міцеліальна стадії. При пророщуванні базидіоспор на штучному поживному середовищі N. sebacea формує білуваті дріжджоподібні колонії (Sampaio et al., 1999;Bauer et al., 2006;Giraldo, 2019). Оскільки гриб не синтезує каротиноїдів, він є низькотолерантним до світла (Libkind et al., 2011). ...
The article provides information about a rare fungicolous fungus Naohidea sebacea first recorded in Ukraine in Roztochchia Nature Reserve. It was found to parasitize Dothiorella iberica (Botryosphaeriaceae, Botryosphaeriales, Dothideomycetes, Ascomycota). This host is reported for N. sebacea for the first time. Light microscopy, cultural studies, and nucleotide sequence analysis of the ITS-region of the ribosomal DNA were used to identify specimens. The resulting sequences were uploaded to the GenBank database. Based on the analysis of literature sources and personal data, information on morphological features, distribution, and substrate specialization of the fungus is provided.
... Smut fungi present a high diversity of ultrastructural characteristics of septal pores and host plant interaction structures, reflecting complex systematic relationships (Bauer et al., 2001). These micromorphological features, alongside macromorphological and molecular characters, justify the current classification of smut fungi into different orders (Bauer et al., 2001(Bauer et al., , 2006Begerow et al., 2018). ...

... Our study shows that it is not uncommon in house dust in Korea. Cystobasidium is known to be mycoparasitic (Bauer et al. 2006;Wang et al. 2015). One hypothesis that Cystobasidium was positively related with HDMs may be due to its mycoparasitism. ...




House dust mites (HDMs) including Dermatophagoides spp. are an important cause of respiratory allergies. However, their relationship with microorganisms in house dust has not been fully elucidated. Here, we characterized bacteria and fungi associated with HDMs in house dust samples collected in 107 homes in Korea by using DNA barcode sequencing of bacterial 16S rRNA gene, fungal internal transcribed spacer 2 (ITS2) region, and arthropod cytochrome c oxidase I (COI) gene. Our inter-kingdom co-occurrence network analysis and/or indicator species analysis identified that HDMs were positively related with a xerophilic fungus Wallemia, mycoparasitic fungi such as Cystobasidium, and some human skin-related bacterial and fungal genera, and they were negatively related with the hygrophilous fungus Cephalotrichum. Overall, our study has succeeded in adding novel insights into HDM-related bacteria and fungi in the house dust ecosystem, and in confirming the historically recognized fact that HDMs are associated with xerophilic fungi such as Wallemia. Understanding the microbial ecology in house dust is thought to be important for elucidating the etiology of human diseases including allergies, and our study revealed baseline information of house dust ecology in relation to HDMs. The findings could be useful from a perspective of human health.
... Barley is infected by four different rusts, i.e., stem, leaf, yellow, and crown rust, all caused by members of the genus Puccinia, family Pucciniaceae, order Pucciniales, class Pucciniomycetes, subphylum Pucciniomycotina, Phylum Basidiomycota, kingdom Fungi, and domain Eukarya(Bauer et al. 2006). Black stem rust of barley caused by Puccinia graminis f. sp. ...

Barley is regarded as the globe’s fourth major cereal crop. A variety of airborne, seedborne, and soilborne infective agents attack barley, causing a variety of barley diseases and substantial losses in agricultural output. Brown and yellow rusts, smut, net blotches, spot blotches, barley yellow dwarf, and molya disease are among the most serious diseases. In general, employing integrated disease management approaches is the best way to handle barley diseases. Growing resistant or tolerant varieties with the fewest foliar fungicides is the most effective approach for barley disease treatments. However, managing soilborne pathogens in barley plants is problematic due to a deficiency in distinguishing symptoms for diagnosis and the absence of fungicides or nematicides that are effective for these pathogens. Recently, nanotechnology has driven the advancement of creative concepts and agricultural productivity with a broad scope for managing plant infections and pests. The antimicrobial properties of metallic and metal oxide nanoparticulates such as silver, selenium, titanium dioxide, zinc oxide, and iron oxide have been extensively researched. In this chapter, we go over barley disease and the role of nanomaterials in reducing the incidence of disease and diagnosis, as well as barley seed germination, physiology, and nutritional quality of barley grain.KeywordsLeaf rust diseaseNet Blotch diseasePowdery mildewBarley yellow dwarfBarley smutSpot blotchFungicidesNanoparticulate