Figure 2 - uploaded by Sinlapachai Senarat
Content may be subject to copyright.

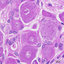
Schematic diagram, micrograph of histology and histochemistry of atretic follicles in Rastrelliger brachysoma ; (A-B) atretic follicle of previtellogenic stage (Ap), A = 100 m, B = 50 m; (D-I) Normal vitellogenic stage (V) = 70 m; (C, K-O) Stage Is = 50 m; (J, P-T) Stage II = 50 m; (U-Z) Stage III = 50 m; (Z1, a-e) Stage IV = 50 m; (Z2, f-j) Stage V = 50 m; Apt = apoptosis, B = basement membrane, Bv = blood vessel, Ca = Cortical alveoli, G = granulosa cell, Hg = hypertrophy of granulosa cell, Lc = lymphocytes, Ly = liquefaction of yolk granules, N = nucleus, P = previtellogenic stage, Pt = pynotic nuclei, T = theca cell, VC = vacuole, Wbc = white blood cells, Y = yolk granules, Zi = inner layer of zonapellucida, Zo = outer layer of zonapellucida, Zp = zonapellucida, * = degeneration of yolk granule, ** = degeneration of cortical alveoli.(MT = Masson’s trichrome, PAS = periodic acid-schiff, AB = aniline blue, Rt = reticulin method).
Source publication






An understanding of atretic follicles in wild population is required before investigating the reproductive cycle and
spawning season but these follicles have never been reported on adult short mackerel Rastrelliger brachysoma (Bleeker,
1851).
Contexts in source publication
Context 1
... characterization of atretic follicle in the pre- vitellogenic stage was similarly seen with normal stage, but the ooplasm shown as basophilic cytoplasm with a surrounding thin follicular layer (PAS staining) (Figure 2A and 2B). ...
Context 2
... normal vitellogenic stage, histo- logical details of several atretic follicles during vitellogenic stage in this species were detected. It could be completely divided into four steps according to shape, characterizations of nucleus and follicle complex ( Figure 2D to 2I). ...
Context 3
... in shape and degeneration with separation between inner and outer layers of the zonapellucida were initially and continuously detected. Additionally, hypertro- phies together with pyknosis of some granulosa and theca cells were detected ( Figure 2C and 2K). Based on histo- chemistry, the cortical alveoli were still positive with MT as reddish and AB as bluish. ...
Context 4
... on histo- chemistry, the cortical alveoli were still positive with MT as reddish and AB as bluish. Other characterizations, yolk granules with slight positively stain with PAS reaction, were specially surrounded by basement membrane (black line with RT) ( Figure 2L to 2O). Stage II, the early stage of stage II was irregular in shape. ...
Context 5
... hypertrophy of granulosa cells were also continuously proliferated, which each cell had a spherical nucleus with surrounding the eosinophilic cytoplasm. Externally, the thecal cell layer was located around few blood vessels ( Figure 2J and 2P). AB, MT and PAS reactions conformed to previous stage but RT reaction distinctly showed the degeneration due to the fragment of basement membrane ( Figure 2Q to 2T). ...
Context 6
... the thecal cell layer was located around few blood vessels ( Figure 2J and 2P). AB, MT and PAS reactions conformed to previous stage but RT reaction distinctly showed the degeneration due to the fragment of basement membrane ( Figure 2Q to 2T). ...
Context 7
... granulosa and theca were not separated and rarely seen. Surprisingly, in this stage, the leucocytes were mostly found and continued to be found near oocyte ( Figure 2U and 2V). Histochemically, the cortical alveoli were slightly seen and began to degenerate (AB and MT stains) whereas RT reaction confirmed that no basement membrane was seen during this stage ( Figure 2W to 2Z). ...
Context 8
... in this stage, the leucocytes were mostly found and continued to be found near oocyte ( Figure 2U and 2V). Histochemically, the cortical alveoli were slightly seen and began to degenerate (AB and MT stains) whereas RT reaction confirmed that no basement membrane was seen during this stage ( Figure 2W to 2Z). ...
Context 9
... the follicular cells became phagocytize degenerating materials because several leucocytes were presented among a few blood vessels ( Figure 2Z1 and 2a). No mucopolysaccharide was observed according to AB and MT, indicating the complete degeneration of cortical alveoli (Figure 2b and 2e). ...
Context 10
... follicle (both yolk granules and zona pellucida) itself was completely digested. The large vacuole associated with yellow-brownish pigments within the ooplasm was accumulated ( Figure 2Z2 and 2f). Some follicular cells were also presented. ...
Context 11
... follicular cells were also presented. Moreover, this stage was surrounded by fibroblast-like cells, as strongly positive with MT and PAS stained (Figure 2g and 2h). It should be noted that this stage could potentially cause by the apoptosis under RT reaction (Figure 2j). ...
Context 12
... this stage was surrounded by fibroblast-like cells, as strongly positive with MT and PAS stained (Figure 2g and 2h). It should be noted that this stage could potentially cause by the apoptosis under RT reaction (Figure 2j). The leucocytes were found around the apoptotic follicles (based on RT stained). ...
Citations





We investigated the digestive biology of two prevalent leiognathid species in Pranburi River estuary, Thailand: the decorated ponyfish (Nuchequula gerreoides) and the splendid polyfish (Eubleekeria splendens). A total of 632 samples collected from February to April and September to November 2017 were analysed using morphological and histological approaches. The overall structures were similar between the species: a short mucous‐cell‐rich oesophagus region, a well‐developed gastric gland uniformly present across the stomach's mucosal layer, and three finger‐like pyloric caeca between the stomach and intestine. However, there were marked differences in the mouth, gill raker, and intestinal coefficient (IC). N. gerreoides had a relatively longer mouth, smoother gill rakers, and an IC of 1.08 ± 0.01, similar to those of other carnivorous fish. In contrast, the gill raker of E. splendens had more villiform teeth that can filter‐feed better, and their IC was 2.16 ± 0.02 (i.e., longer intestine). Although digestive structures were generally similar between the ponyfishes, these differences suggest that N. gerreoides is relatively carnivorous with stronger suction, whereas E. splendens may be an omnivorous or herbivorous filter‐feeder.


The scalloped perchlet Ambassis nalua is one of the dominant fishes in the Estuarine Pranburi River, Thailand. It is suggested that this fish is in the secondary trophic level with a carnivorous nature. Studies on digestive system will help us further identify the niche of this species in the food web/food chain. The present study therefore aimed to report the detailed structure and ultrastructure of A. nalua digestive system. Fish samples (n = 30) with a total length of 5.7 ± 0.5 cm were obtained using beach seines from the Estuarine Pranburi River. Their digestive tract length and intestine coeficient were 3.6 ± 0.07 cm and 0.91, respectively. Light microscopic observation showed that the digestive wall comprised four layers, namely mucosa, submucosa, muscularis, and serosa. The prominent mucous‐secreting cells were found in the mucosal oesophagus. The stomach had many gastric folds, with height and width being 649.76 ± 85.15 and 370.30 ± 68.56 μm, respectively. Gastric glands were found in the anterior stomach but not in the posterior stomach. Each gastric gland was made up of a single type of columnar cells. The gastric cells were ultrastructurally characterized by numerous mitochondria and well‐developed secretory granules of varying sizes. A few small vacuoles were also identified in the apical area of the gastric cells. The intestine had two regions (anterior and posterior intestines), and pyloric caecum was absent. The density of the goblet cell was significantly higher in the posterior intestine. These results provide basic knowledge of the digestive system of A. nalua, and the low intestine coefficient and the absence of pyloric caecum suggest the carnivorous feeding habit of this species.