Figure 2 - uploaded by Barbara Ghislain
Content may be subject to copyright.
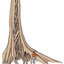
Patterns of lignification of the G-layers in tension wood. Tension wood (left) and opposite wood (right) of Hevea guianensis (A), Macrolobium bifolium (B), Protium guianense (C), Manilkara bidentata (D), Guarea guidonia (E), Licania macrophylla (F), Sextonia rubra (G), Eperua grandiflora (H), Lacistema grandifolium (I), Tachigali melinonii (J). G-layers are either unlignified (A-C), lignified (D-F) or intermediate (G-J). Cambium is located on the right.
Source publication








Angiosperm trees produce tension wood to actively control their vertical position. Tension wood has often been characterised by the presence of an unlignified inner fibre wall layer called the G-layer. Using this definition, previous reports indicate that only one-third of all tree species have tension wood with G-layers. Here we aim to (i) describ...
Contexts in source publication
Context 1
... and one group for reaction wood without G-layers. For tension wood with G-layers, we defined three anatomical groups based on the main trend in overall G-layer lignification of wood for which maturation is completed, including late lignification. If most of the mature G-layers are unlignified, the anatomical group is labeled "G-layer unlignified" (Fig. 2 A-C). If mature G-layers have unlignified as well as lignified G-layers or have G-layers not completely lignified, the anatomical group is referred to as "G-layer partially lignified" (Fig. 2 G-J). If all mature G-layers are lignified, the anatomical group is named "G-layer lignified" (Fig. 2 D-F). The cell maturation phase could not always ...
Context 2
... maturation is completed, including late lignification. If most of the mature G-layers are unlignified, the anatomical group is labeled "G-layer unlignified" (Fig. 2 A-C). If mature G-layers have unlignified as well as lignified G-layers or have G-layers not completely lignified, the anatomical group is referred to as "G-layer partially lignified" (Fig. 2 G-J). If all mature G-layers are lignified, the anatomical group is named "G-layer lignified" (Fig. 2 D-F). The cell maturation phase could not always be observed, either because the tree was not growing at the sampling date (all cells adjacent to the cambium were mature) or in a few cases because the tension wood sector was located far ...
Context 3
... the anatomical group is labeled "G-layer unlignified" (Fig. 2 A-C). If mature G-layers have unlignified as well as lignified G-layers or have G-layers not completely lignified, the anatomical group is referred to as "G-layer partially lignified" (Fig. 2 G-J). If all mature G-layers are lignified, the anatomical group is named "G-layer lignified" (Fig. 2 D-F). The cell maturation phase could not always be observed, either because the tree was not growing at the sampling date (all cells adjacent to the cambium were mature) or in a few cases because the tension wood sector was located far from the cambium in some naturally tilted juvenile trees. When no unlignified G-layer was observed and ...
Context 4
... G-layers appear in tension wood of most species, mostly near the cambium. However, species in which the G-layers remain unlignified far from the cambium, as in Populus ( Pilate et al. 2004), are quite rare. Far from the cambium, unlignified G-layers are observed in only 9 species (Fig. 2A). For 12 species G-layers are mainly unlignified but slight staining for lignin is sometimes observed inside fibres, either towards the edges of the cell (Fig. 2B) or in the whole ...
Context 5
... the G-layers remain unlignified far from the cambium, as in Populus ( Pilate et al. 2004), are quite rare. Far from the cambium, unlignified G-layers are observed in only 9 species (Fig. 2A). For 12 species G-layers are mainly unlignified but slight staining for lignin is sometimes observed inside fibres, either towards the edges of the cell (Fig. 2B) or in the whole ...
Context 6
... the cell level, the G-layer has either a homogeneous or heterogeneous staining of the whole G-layer, which can be more or less intense (Fig. 1). At the tissue level, in 57% of the species all G-layers were fully and homogeneously lignified far away from the cambium (Fig. 2D-F). In 19% of the species, lignification could occur in some G-layers and not in others, with heterogeneous staining or not and in a random pattern or not (Fig. 2 G-J). For example, in Pourouma sp.1, lignified G-layers occurred on the borders of the tension wood arc ...
Context 7
... the whole G-layer, which can be more or less intense (Fig. 1). At the tissue level, in 57% of the species all G-layers were fully and homogeneously lignified far away from the cambium (Fig. 2D-F). In 19% of the species, lignification could occur in some G-layers and not in others, with heterogeneous staining or not and in a random pattern or not (Fig. 2 G-J). For example, in Pourouma sp.1, lignified G-layers occurred on the borders of the tension wood arc ...
Context 8
... and one group for reaction wood without G-layers. For tension wood with G-layers, we defined three anatomical groups based on the main trend in overall G-layer lignification of wood for which maturation is completed, including late lignification. If most of the mature G-layers are unlignified, the anatomical group is labeled "G-layer unlignified" (Fig. 2 A-C). If mature G-layers have unlignified as well as lignified G-layers or have G-layers not completely lignified, the anatomical group is referred to as "G-layer partially lignified" (Fig. 2 G-J). If all mature G-layers are lignified, the anatomical group is named "G-layer lignified" (Fig. 2 D-F). The cell maturation phase could not always ...
Context 9
... maturation is completed, including late lignification. If most of the mature G-layers are unlignified, the anatomical group is labeled "G-layer unlignified" (Fig. 2 A-C). If mature G-layers have unlignified as well as lignified G-layers or have G-layers not completely lignified, the anatomical group is referred to as "G-layer partially lignified" (Fig. 2 G-J). If all mature G-layers are lignified, the anatomical group is named "G-layer lignified" (Fig. 2 D-F). The cell maturation phase could not always be observed, either because the tree was not growing at the sampling date (all cells adjacent to the cambium were mature) or in a few cases because the tension wood sector was located far ...
Context 10
... the anatomical group is labeled "G-layer unlignified" (Fig. 2 A-C). If mature G-layers have unlignified as well as lignified G-layers or have G-layers not completely lignified, the anatomical group is referred to as "G-layer partially lignified" (Fig. 2 G-J). If all mature G-layers are lignified, the anatomical group is named "G-layer lignified" (Fig. 2 D-F). The cell maturation phase could not always be observed, either because the tree was not growing at the sampling date (all cells adjacent to the cambium were mature) or in a few cases because the tension wood sector was located far from the cambium in some naturally tilted juvenile trees. When no unlignified G-layer was observed and ...
Context 11
... G-layers appear in tension wood of most species, mostly near the cambium. However, species in which the G-layers remain unlignified far from the cambium, as in Populus ( Pilate et al. 2004), are quite rare. Far from the cambium, unlignified G-layers are observed in only 9 species (Fig. 2A). For 12 species G-layers are mainly unlignified but slight staining for lignin is sometimes observed inside fibres, either towards the edges of the cell (Fig. 2B) or in the whole ...
Context 12
... the G-layers remain unlignified far from the cambium, as in Populus ( Pilate et al. 2004), are quite rare. Far from the cambium, unlignified G-layers are observed in only 9 species (Fig. 2A). For 12 species G-layers are mainly unlignified but slight staining for lignin is sometimes observed inside fibres, either towards the edges of the cell (Fig. 2B) or in the whole ...
Context 13
... the cell level, the G-layer has either a homogeneous or heterogeneous staining of the whole G-layer, which can be more or less intense (Fig. 1). At the tissue level, in 57% of the species all G-layers were fully and homogeneously lignified far away from the cambium (Fig. 2D-F). In 19% of the species, lignification could occur in some G-layers and not in others, with heterogeneous staining or not and in a random pattern or not (Fig. 2 G-J). For example, in Pourouma sp.1, lignified G-layers occurred on the borders of the tension wood arc ...
Context 14
... the whole G-layer, which can be more or less intense (Fig. 1). At the tissue level, in 57% of the species all G-layers were fully and homogeneously lignified far away from the cambium (Fig. 2D-F). In 19% of the species, lignification could occur in some G-layers and not in others, with heterogeneous staining or not and in a random pattern or not (Fig. 2 G-J). For example, in Pourouma sp.1, lignified G-layers occurred on the borders of the tension wood arc ...
Similar publications









Environmental conditions can change both the quantity and quality of wood formation. This study aimed to evaluate anatomical and chemical changes in the wood of a Eucalyptus urophylla S.T. Blake clone cultivated in four sites of wide climatic conditions in Brazil. Radial samples were used to evaluate xylem anatomy along the growth cycles. Samples w...
Citations
... Further, Onaka [28] found gelatinous fibers in only 40% of the tension wood of 346 Japanese tree species. In addition, some tension wood fiber cells exhibit a multilayer structure with alternating thick and thin layers (Figure 5a,b), and the number of layers in their secondary cell wall is positively correlated with the severity of the tension wood [20,21,45,46]. Moreover, the S 2 layer of cells in the tension wood without G-layers also exhibits some structural features similar to G-layers, such as small microfibrillar angles, large cellulose crystallite sizes, and increased crystallinity [9,47,48]. ...
... Further, Onaka [28] found gelatinous fibers in only 40% of the tension wood of 346 Japanese tree species. In addition, some tension wood fiber cells exhibit a multilayer structure with alternating thick and thin layers (Figure 5a,b), and the number of layers in their secondary cell wall is positively correlated with the severity of the tension wood [20,21,45,46]. Moreover, the S2 layer of cells in the tension wood without G-layers also exhibits some structural features similar to Glayers, such as small microfibrillar angles, large cellulose crystallite sizes, and increased crystallinity [9,47,48]. ...
... However, due to a lack of understanding of the diversity of tension wood at that time, it was not comprehensive to judge whether it was reaction tissue based solely on the existence of the gelatinous layer. Recent studies have found that some broadleaved trees have tension fibers with a multi-layered and alternating thin and thick cell wall structure similar to that found in bamboo fibers (Figure 5c), without the presence of a gelatinous layer [45,46]. Moreover, the arrangement of microfibrils in the cell walls of tension wood are almost parallel to the main axis, just like in bamboo fibers. ...
To maintain or adjust posture under the challenges of gravity and increased self-weight, or the effects of light, snow, and slope, plants have the ability to develop a special type of tissue called reaction tissue. The formation of reaction tissue is a result of plant evolution and adaptation. The identification and study of plant reaction tissue are of great significance for understanding the systematics and evolution of plants, the processing and utilization of plant-based materials, and the exploration of new biomimetic materials and biological templates. Trees’ reaction tissues have been studied for many years, and recently, many new findings regarding these tissues have been reported. However, reaction tissue requires further detailed exploration, particularly due to their complex and diverse nature. Moreover, the reaction tissues in gymnosperms, vines, herbs, etc., which display unique biomechanical behavior, have also garnered the attention of research. After summarizing the existing literature, this paper provides an outline of the reaction tissues in woody plants and non-woody plants, and lays emphasis on alternations in the cell wall structure of the xylem in softwood and hardwood. The purpose of this paper is to provide a reference for the further exploration and study of reaction tissues with great diversity.
... Although the absolute changes in lignin content that are visualised by the double-staining method remain unknown, there is evidence for qualitative similarities between double-staining, autofluorescence imaging and Raman spectroscopy 37,58 . In contrast to polarised light microscopy that does not visualise lignin, the staining with Safranin and Astra Blue can reveal different degrees of lignin content in wood cell walls 57 . Thin sections were washed with water and dehydrated with ethanol at increasing concentration, finally embedded in Canada balsam under the cover glass. ...




More than 200 years after von Humboldt’s pioneering work on the treeline, our understanding of the cold distribution limit of upright plant growth is still incomplete. Here, we use wood anatomical techniques to estimate the degree of stem cell wall lignification in 1770 plant species from six continents. Contrary to the frequent belief that small plants are less lignified, we show that cell wall lignification in ‘woody’ herbs varies considerably. Although trees and shrubs always exhibit lignified cell walls in their upright stems, small plants above the treeline may contain less lignin. Our findings suggest that extremely cold growing season temperatures can reduce the ability of plants to lignify their secondary cell walls. Corroborating experimental and observational evidence, this study proposes to revisit existing theories about the thermal distribution limit of upright plant growth and to consider biochemical and biomechanical factors for explaining the global treeline position.
... Fibres containing a gelatinous layer are widespread in the plant kingdom and can be present in various organs and tissues . Depending on their cell wall composition, fibres can be classified either as lignin-rich (i.e., wood fibres) or virtually lignin-free, such as part of tension wood fibres (Ghislain et al., 2019) or bast fibres found in flax, hemp, ramie, nettle, etc (Gorshkova and Morvan, 2006). It is this latter category of fibres that is often named gelatinous fibre. ...



Trees generate mechanical stresses at periphery of stem and branches to improve their strength and to control the orientation of their axes. This key factor in the biomechanical design of trees, named "maturation stress", occurs in wood fibres during cellular matu-ration when their secondary cell wall thickens. In this study, the spatial and temporal stiffening kinetics of the different cell wall layers were recorded during fibre matura-tion on a sample of poplar tension wood using atomic force microscopy. The thickening of the different layers was also recorded. The stiffening of the CML, S1 and S2-layers was initially synchronous with the thickening of the S2-layer and continued a little after the S2-layer reached its final thickness as the G-layer began to develop. In contrast, the global stiffness of the G-layer, which initially increased with its thickening, was close to stable long before it reached its final maximum thickness. A limited radial gradient of stiffness was observed in the G-layer, but it decreased sharply on the lumen side, where the new sub-layers are deposited during cell wall thickening. Although very similar at the ultrastructural and biochemical levels, the stiffening kinetics of the poplar G-layer appears to be very different from that described in maturing bast fibres.
... Fibres containing a gelatinous layer are widespread in the plant kingdom and can be present in various organs and tissues . Depending on their cell wall composition, fibres can be classified either as lignin-rich (i.e., wood fibres) or virtually lignin-free, such as part of tension wood fibres (Ghislain et al., 2019) or bast fibres found in flax, hemp, ramie, nettle, etc (Gorshkova and Morvan, 2006). It is this latter category of fibres that is often named gelatinous fibre. ...

... Rods for properties measurement OW sector LW sector Figure 12. Comparative anatomy of compression wood and normal wood for the conifer species CW: compression wood; NW: normal wood For hardwoods, a majority of G-layer type TW (3 poplars, Miconia, Carapa, Ocotea, Cecropia, Eperua) were studied (Table 3), two species had a lignified G layer (Eschweilera & Qualea), one a peculiar multilayered G layer (Laetia) and three no G layer fibre (Jacaranda, Virola, Simarouba), according to a recent classification based on 242 tropical species (Ghislain et al 2019). Measurement of MFA on the 3 species represented in Fig. 13 (Ruelle et al 2007) showed lower values for TW (2° to 14°) than for NW (10° to 35°), with some overlap around 10°-14°. ...
Living wood in the tree performs a muscular action by generating forces at the sapwood periphery and residual strains in dead sapwood fibres. Dissymmetric force generation around the tree trunk is the motor system allowing movement, posture control and tree reshaping after accidents. Rather young trees are able to restore the verticality of their trunk after accidental rotation of the soil-root system due to wind or landslide, leading to typically curved stems shape. The very high dissymmetry of forces for the motor action is associated with the occurrence of reaction wood on one side of the inclined stem during many successive years. A method to reconstitute this biomechanical history from observations after tree felling, on either green or dry wood, is discussed. A selection of 17 trees from 15 different species (13 different families), tropical and temperate, hardwoods and softwoods, were selected and peripheral residual strains were measured in situ before felling, on 8 positions for each stem. Matched wooden rods were sawn and measured for their mechanical and physical properties in the green and dry states, allowing the estimation of tree growth stress, i.e., the force created by the living wood. It was possible to build easy-to-use conversion coefficients between the growth stress indicator (GSI), measured in situ by the single hole method, and growth strain and growth stress with the knowledge of basic density and green longitudinal elastic modulus. Maturation strain, specific modulus (as a proxy of micro-fibril angle) and longitudinal shrinkage are properties independent from basic density, whose variation among species was very large. For the whole range of compression wood, normal wood and tension wood, strong relationships between these 3 properties were observed, but together no single model, based on cell-wall microfibril angle only, could be defined. Growth forces are the product of 4 parameters: ring width, basic density, basic specific modulus and maturation strain, all of them being the result of wood formation. Thanks to the wide range of wood types and species, simple and highly significant formulas were obtained for the relationship between basic and dry density, green and dry longitudinal modulus of elasticity, basic and dry specific modulus. To estimate ring width in the green state from values in dry state, radial shrinkage needs to be measured afterwards on dry specimens. Maturation strains is less accurately linked to late measurements on dry wood, but longitudinal shrinkage offers a rather good solution for an estimation provided that the wood type (softwood, hardwood with-G layer, hardwood without G-Layer) is known.
... G-layers are largely composed of crystalline cellulose with a lower lignin content than that in SCW (Onaka, 1949;Timell, 1969), although lignification of G-layers is found in some tree species (Ghislain and Clair, 2017;Ghislain et al., 2019;Higaki et al., 2017;Roussel and Clair, 2015). The contents of galactan and xyloglucan, two non-cellulosic polysaccharides, are increased in G-layers, but xylan content is decreased in many tree species (Baba et al., 2009;Gorshkova et al., 2015;Guedes et al., 2017;Kim and Daniel, 2019;Nishikubo et al., 2007). ...
... GF, G-fiber; f, wood fiber without SCW. Ghislain and Clair, 2017;Ghislain et al., 2019). G-layers are formed on the inner side of the SCW in wood fibers. ...
... For example, wood fibers with or without a G-layer are differentiated side by side in Hevea brasiliensis and Populus euramericana (Araki et al., 1982;Nobuchi et al., 2011). Similarly, fibers with lignified Glayers occur next to fibers with unlignified G-layers in several hardwood species (Clair et al., 2006;Ghislain et al., 2019;Hirohashi, et al., 2012;Roussel and Clair, 2015). These results suggest that a differentiating single wood fiber recognizes its developmental stage and responds to cellular signals at a specific developmental stage prior to G-layer formation regardless of the cellular conditions of the surrounding wood fibers. ...




Cell walls, especially secondary cell walls (SCW), maintain cell shape and reinforce wood, but their structure and shape can be altered in response to gravity. In hardwood trees, tension wood forms along the upper side of a bending stem and contains wood fiber cells that have a gelatinous layer (G-layer) inside the SCW. In a previous study, we generated nst/snd quadruple-knockout aspens, in which SCW formation was impaired in 99% of the wood fiber cells. In this study, we produced nst/snd triple-knockout aspens, in which a large number of wood fibers had thinner SCW than the wild-type (WT) and some had no SCW. Because SCW layers are always formed prior to G-layer deposition, the nst/snd mutants raise interesting questions of whether the mutants can form G-layers without SCW and whether they can control their postures in response to changes in gravitational direction. The nst/snd mutants and the WT plants showed growth eccentricity and vessel frequency reduction when grown on an incline, but the triple mutants recovered their upright growth only slightly, and the quadruple mutants were unable to maintain their postures. The mutants clearly showed that the G-layers were formed in SCW-containing wood fibers but not in those lacking the SCW. Our results indicate that SCW are essential for G-layer formation and posture control. Furthermore, each wood fiber cell may be able to recognize its cell wall developmental stage to initiate the formation of the G-layer as a response to gravistimulation.
... It is evident that the lack of cell wall lignification is always stronger in BRs compared to pBRs. However, in both cases, we can assume that the bluer the cell walls, the less lignified they are (Gerlach 1984, Piermattei et al 2015, Ghislan et al 2019, Crivellaro and Büntgen 2020, Tardif et al 2020. ...


Blue Rings' (BRs) are distinct wood anatomical anomalies recently discovered in several tree species from different sites. While it is evident that they are associated with a cooling-induced lack of cell wall lignification, BRs have yet to be evaluated systematically in paleoclimate studies. Here, we present a continuous wood anatomical assessment of 31 living and relict pine samples from a high-elevation site in the central Spanish Pyrenees that span the period 1150-2017 CE at annual resolution. While most BR years coincide with cold summer temperatures and many BRs follow large volcanic eruptions, some were formed during overall warm summers. We also see a differential response between eruptions: the Samalas eruption is followed by 80% BRs in 1258, but only a modest signal is evident after the 1815 Tambora eruption, and there are no wood anatomical effects of the Laki eruption in 1783-1784. Apparently linked to a cluster of tropical eruptions in 1695 and 1696 CE, 85% BRs occurred in 1698. This new wood anatomical evidence is corroborated by the record of sulphur deposition in polar ice cores, and corresponds with catastrophic famine and unprecedented mortality in Scotland. The extremely rare occurrence of consecutive BRs in 1345 and 1346 marks the onset and spread of the Black Death, Europe's most devastating plague pandemic. In their ability to capture severe ephemeral cold spells, as short as several days or weeks, BR chronologies can help to investigate and understand the impacts of volcanism on climate and society.
In this book, Lee A. Newsom offers an overview of wood in archaeology --how and where it is preserved and analyzed, its relevance to paleoecological and paleoenvironmental questions, as well as its role as an important source of information in modern archaeological science and related historical disciplines. Her book addresses a range of questions about wood reliance practices, sustainability, and the overall relevance of forest ecosystems to past cultures and cultural evolution. Newsom provides a step-by-step treatment of archaeological analysis with clear explanations and examples from various corners of the world. She also shows how the study of archaeological wood is relevant to modern restoration ecology and conservation biology that tracks long-term ancient ecosystems, including questions of global change. Demonstrating the vital role of wood and timber resources to past human societies, her book will interest scholars and students of archaeology, historical ecology, paleoecology, and wood science.


Gelatinous fibers (G-fibers) are specialized contractile cells found in a diversity of vascular plant tissues, where they provide mechanical support and/or facilitate plant mobility. G-fibers are distinct from typical fibers by the presence of an innermost thickened G-layer, comprised mainly of axially oriented cellulose microfibrils. Despite the disparate developmental origins—tension wood fibers from the vascular cambium or primary phloem fibers from the procambium—G-fiber development, composition, and molecular signatures are remarkably similar; however, important distinctions do exist. Here, we synthesize current knowledge of the phylogenetic diversity, compositional makeup, and the molecular profiles that characterize G-fiber development and highlight open questions for future investigation.
Many plant movements are facilitated by contractile cells called gelatinous fibers (G‐fibers), but how G‐fibers function in the climbing movements of woody vines remains underexplored. In this Insight, we compare the presence and distribution of G‐fibers in the stems of stem‐twiners, which wrap around supports, with non‐stem‐twiners, which attach to supports via tendrils or adventitious roots. An examination of 164 species spanning the vascular plant phylogeny reveals that G‐fibers are common in stem‐twiners but scarce in non‐stem‐twiners, suggesting that G‐fibers are preferentially formed in the organ responsible for movement. When present, G‐fibers are in the xylem, phloem, pericycle, and/or cortex. We discuss the hypothesis that G‐fibers are foundational to plant movement and highlight research opportunities concerning G‐fiber development and function.