Figure 5 - uploaded by Patrick Lestienne
Content may be subject to copyright.

Mitochondrial respiratory chain. For mammals, the respiratory chain consists of four enzyme complexes (complexes I – IV) and two intermediary substrates (coenzyme Q and cytochrome c). The NADH+H + and FADH 2 produced by the intermediate metabolism are oxidized further by the mitochondrial respiratory chain to establish an electrochemical gradient of protons, which is finally used by the F1F0-ATP synthase (complex V) to produce ATP, the only form of energy used by the cell. In this simple representation of the respiratory chain, the supramolecular organization (supercomplexes, dimers) is not shown.
Source publication







In this review, we discuss the concept of metabolic remodeling and signaling in tumors, specifically the various metabolites that participate in the regulation of gene expression in cancer cells. In particular, pyruvate, oxaloacetate, succinate and fumarate, four mitochondrial metabolites, activate genes relevant for tumor progression. When the bal...
Context in source publication
Context 1
... the inner membrane and the ultimate phosphorylation of ADP by the F F -ATP synthase (complex V). In 1963, mtDNA (Figure 1) was characterized by Nass and Schatz and then entirely sequenced by Sanger and co-workers in 1981. By that time, L. Margulis had proposed the endosymbiotic theory of mitochondrial evolution, which states that aerobic bacteria, upon symbiosis, transferred most of their DNA to the nucleus of the protocells and gained the selective advantage of being able to consume oxygen while producing energy in the form of ATP. By examining the polymorphisms in mtDNA, H. Blanc discovered its maternal inheritance; others discovered that it had a mutation rate 10 times higher than that of nuclear DNA. Furthermore, some mitochondrial DNA genes were found to be localized in the chromosomes, but are silent. In 1979, Barell found differences in the genetic code as compared with the universal one. Two years later, the entire sequence of the human mitochondrial DNA was established by F. Sanger and collaborators. The first diseases associated with mtDNA deletions were reported in early 1988; these disorders are associated with several clinical symptoms, such as myopathy, ptosis, external ophthalmoplegia and cardiac block branch (for an extensive review, see (2)). The first point mutation identified was associated with Leber’s disease, which is mainly characterized by visual loss. During the 1990s, the three- dimensional structures of the cytochrome c oxidase (complex IV) and of the ATP synthase were determined by the groups of S. Yoshikawa and J. Walker, respectively. The identification of proapoptotic factors that are released by this organelle indicated that mitochondria play a pivotal role in deciding cell survival or death through intracellular signaling. Apoptosis can be divided in three phases: induction, execution and degradation (Figure 2). In this review, we will not cover the downregulation of mitochondrial apoptotic mechanisms in cancer cells. We will focus more on the particularities of their energy metabolism in relationship with metabolic signaling and tumor biology. In the last decade, much progress has been made in deciphering the link between mitochondrial structure and function with the discovery (consideration) of a networked and dynamic organization of the mitochondrion in living human cells of diverse origin (Figure 3A). Studies examining the mitochondrial interior by electron tomography helped to define the "cristae-junction model" (3), which separates the inner boundary membrane (IBM) closely opposed to the outer membrane from the cristae membrane that projects into the matrix compartment (Figure 3B). These two components may be connected by tubular structures called cristae junctions of relatively uniform size. Recent advances in mitochondrial research revealed that morphological changes of the mitochondria, both overall organization and internal structure, are intimately linked with bioenergetics and apoptosis (for reviews, see (4) and (5), respectively). The additional discovery of pathogenic mutations in genes that are essential for fusion and fission of the mitochondrial network has added complexity to the physiopathological mechanisms of these diseases and the regulation of organellar functions as diverse as energetics, signaling, biosyntheses or apoptosis (6). Prior to discussing the abnormalities of cancer cells energetics it is necessary to give a brief introduction on the metabolic pathways by which glucose is converted in ATP, the only form of energy considered by the cell (Figure 4). Briefly, glucose enters the cells through specific transporters and becomes phosphorylated by hexokinase to G 6 P which is further cleaved in two molecules of glyceraldehyde-3-phosphate and dephosphorylated by the pyruvate kinase in the presence of ADP to yield pyruvate. These reactions produce two molecules of ATP and are collectively known as glycolysis (anaerobic and cytosolic) in contrast with respiration (aerobic and mitochondrial). The end product of glycolysis pyruvate can be transformed in lactate by the lactate dehydrogenase in the presence of an excess of NADH + H + during poor respiration, or it can enter the mitochondria to undergo an oxidative decarboxylation in the presence of coenzymes (NAD + , FAD) which will fuel the respiratory chain to consume oxygen and yield ATP and H 2 O. Inside the mitochondrion, the intermediate metabolism encompasses the Krebs cycle and the beta-oxidation which provide acyl-CoA and acetyl- CoA but also NAD (P)H + H + and FADH 2 . Mitochondria contain other enzymes involved in catabolism (urea cycle) or anabolism with the synthesis of amino acids and uridine. They also play a pivotal role in the phosphorylation of nucleotide diphosphates, notably thymidine. The five complexes of the respiratory chain are embedded within the inner boundary membrane and the cristae membrane (ratio 1 to 6). Only 13 out of the hundred of proteins of the entire OXPHOS system (Figure 5) are encoded by the mtDNA which uses a specific genetic code. The inner membrane is impermeable to protons so that a proton electrochemical gradient can be formed upon the proton extrusion from the matrix to the intermal membrane space by the respiratory chain complexes I, III and IV. According to the chemiosmotic theory of P. Mitchell (7), the resulting electric membrane potential (deltaPsi) and pH difference (deltapH) caused by this process create a strong proton- motive force that is used for the condensation of ADP and Pi by the ATP synthetase to produce ATP. ATP is then exported to the cytoplasm by the adenine nucleotide translocator (ANT) in exchange with cytosolic ADP. Aerobic respiration from glucose produces 38 moles of ATP, which is 19 times more than what glycolysis can produce. However, the rate of ATP production by the OXPHOS system is slower than glycolysis (8). O. Warburg noticed that a significant number of tumors use glycolysis more than respiration to produce the vital ATP, even under normal O 2 pressure. As a result the extracellular medium is acidified. The genetic, biochemical and molecular basis of this phenomenon, which is called the "Warburg effect", still remain puzzling. Its occurrence in all tumors is also challenged by the discovery of cancers that utilize more the mitochondrion to produce ATP. An elevated uptake of glucose (by a factor of 10) is generally observed in human living tumoral tissues by positron- emission tomography (PET) using the isotope analog glucose tracer 2- ( 18 F) fluoro-2-deoxy-D-glucose (FDG). This index was further correlated with increased malignancy and invasiveness (9, 10); for some tumors, a close correlation was observed between the degree of differentiation, growth rate and glucose accumulation (1, 11). Such clinical observations suggest that a large amount of glucose enters tumors, but the contribution of glycolysis and OXPHOS to glucose catabolism cannot be determined with this technique. To do so, bioenergetic studies are required on tissue sample or tumor derived cell lines. In this manner the extent of ATP produced by glycolysis was measured on various cancer cell lines and revealed that Warburg predictions were not systematic. For instance the bioenergetic analysis of two human lung cancer cell lines verified Warburg's theory but also precised that OXPHOS is not inactive per se : it operates at a low-capacity and is repressed by the presence of glucose (Crabtree effect) (12). In striking contrast, an analysis of MCF-7 cells originating from a mammary gland epithelial adenocarcinoma revealed that ATP production is 80 % oxidative in glucose medium (13). Likewise, in hepatoma cells mitochondrial respiration was found to be coupled to ADP phosphorylation and produced 40% of the total cellular ATP in glucose medium (14). More recently, Rodriguez-Enriquez et al. revealed that in HeLa and Hek293 young-spheroids, the contribution of OXPHOS to the total ATP supply was 60% (15). They further studied this feature and evidenced a class of tumor cell lines in which the oxidative metabolism prevails over glycolysis. This situation has been extensively reviewed in (16), and the authors even conclude that "high glycolysis" is not a prerequisite of all cancer cells but could be acquired during the highest proliferative activity and/or in response to stringent micro-environmental conditions, such as intermittent hypoxia and/or glucose limitation. Accordingly, a critical review of numerous studies comparing cancer cells with normal tissues concluded that several tumors derive most of their ATP from mitochondrial oxidative phosphorylation, in striking contrast to Warburg's hypothesis (17). These divergent observations highlight the fundamental question of how variable is the metabolic reprogramming of cancer cells (Figure 6) and what are the relative contributions of oncogenesis, tumor environment, proliferative activity and experimental conditions. The examination of cancer cells metabolic profile should be considered for tumor caracterisation and classification as specific alterations in mitochondrial and glycolytic protein expression were reported in association with changes in cancer cell bioenergetics or apoptosis (18-20). In a large variety of tumors, extensive modifications of the mitochondrion have further been described, including a decrease in organellar biogenesis (11, 21), the alteration of respiratory chain complexes specific activity (22, 23), inhibition of the pyruvate dehydrogenase (PDH) complex (24), truncation of the Krebs cycle with citrate extrusion (25), an increase in the binding of hexokinase II to the mitochondrion (26), variegated changes in organellar shape and size (27) and the accumulation of diverse mutations in mitochondrial DNA (28, 29). More recently, it was proposed that metabolic reprogramming could be used to enhance tumors cells biosynthetic pathway, as required by the deregulated growth and needs. This is ...
Similar publications








The finding that upon neuronal activation glutamate is transported postsynaptically from synaptic clefts and increased lactate availability for neurons suggest that brain mitochondria (BM) utilize a mixture of substrates, namely pyruvate, glutamate, and the tricarboxylic acid cycle metabolites. We studied how glutamate affected oxidative phosphoryl...








mTOR pathway activation and hypervascularity have been identified as important characteristics of pancreatic neuroendocrine tumors (pNETs). Agents targeting angiogenesis and mTOR, such as sunitinib and everolimus (RAD001), have been shown to result in progression-free survival of approximately 11 months in patients with advanced pNETs. Novel treatm...
Citations
... Among the those, this cell line is the most applied for neurodegenerative disorders [8][9][10]. For modeling of neurodegenerative disorders in in vitro, the cell line exposed to 6-Hydroxydopamine (6-OHDA) in many studies [11,12]. ...
... Under intrinsic cellular stresses, such as DNA damage, hypoxia, UV, and exposure to chemicals, mitochondria in neurons release cytochrome c through assembled Bax proteins, and the released cytochromes and activated caspase 9 enhance cellular apoptosis [11]. When 6-OHDA triggers the production of ROS, monoamine oxidase (MAO) A and B are expressed in neurons, and MAOB produces ROS by the degradation of dopamine in neurons [12]. ...
... (www.preprints.org) | NOT PEER-REVIEWED | Posted: 10 May 2024 doi:10.20944/preprints202405.0558.v111 ...


Oxidative neurodegeneration causes various neuronal diseases including Alzhei-14 mer's disease, Parkinson's disease, and Huntington's disease. This study aimed to demonstrate the 15 protective effect of leaf extracts of Allium senescens on 6-hydroxydopamine (6-OHDA)-stressed SH-16 SY5Y cells, known as an optimal cell for neurotoxic research. Levels of apoptotic markers were eval-17 uated using quantitative polymerase chain reaction (qPCR) and flow cytometry. The localization of 18 apoptotic cells in vivo was analyzed using whole-mount immunochemistry and the terminal deox-19 ynucleotidyl transferase dUTP nick end labeling assay. Additionally, reactive oxygen species (ROS) 20 production was estimated using flow cytometry. 6-OHDA induced ROS production in neuroblast 21 cells and in vivo, but treatment with the extract protected against the 6-OHDA-induced increase in 22 ROS levels. Under oxidative stress, the extract performs three protective functions: decreases ROS 23 production, prevents mitochondrial apoptosis, and protects the central and ventral nervous systems. 24 These results also suggest that the extract can be useful for the development of functional foods for 25 the prevention of neural damage due to oxidative stress. 26
... The liver contains a large number of mitochondria, which play an essential role in cellular bioenergetics, regulating redox balance and apoptosis [53]. The mitochondrial respiratory chain consists of five enzymatic complexes, which are mainly responsible for the generation of ATP through oxidative phosphorylation [54]. Oxidative stress can cause mitochondrial dysfunction by disturbing cellular bioenergetics [55]. ...


Silybin (Si) is the main element of silymarin isolated from the seeds of Silybum marianum L. Gaernt., which has superior antioxidant properties. However, the protective role of Si in maintaining liver health under oxidative stress remains ambiguous. This study aimed to investigate the underlying mechanism of the beneficial effect of dietary Si against hepatic oxidative injury induced by paraquat (PQ) in weaned piglets. A total of 24 piglets were randomly allocated to four treatments with six replicates per treatment and 1 piglet per replicate: the control group; Si group; PQ group; and Si + PQ group. Piglets in the control group and PQ group were given a basal diet, while piglets in the Si and Si + PQ groups were given a Si-supplemented diet. On the 18th day, the pigs in the PQ treatment group received an intraperitoneal injection of PQ, and the others were intraperitoneally injected with the same volume of saline. All piglets were sacrificed on day 21 for plasma and liver sample collection. The results showed that dietary Si supplementation mitigated PQ-induced liver damage, as proven by the reduction in liver pathological changes and plasma activity of alanine transaminase and aspartate transaminase. Si also improved superoxide dismutase and glutathione peroxidase activities and total antioxidant capacity, as well as decreased malondialdehyde and hydrogen peroxide concentration in the liver, which were closely related to the activation of the nuclear factor-erythroid 2-related factor 2 signaling pathway. Meanwhile, Si reduced tumor necrosis factor-α and interleukin-8 production and their transcript levels as well as abrogated the overactivation of nuclear factor-κB induced by PQ. Importantly, Si improved mitochondrial function by maintaining mitochondrial energetics and mitochondrial dynamics, which was indicated by the elevated activity of mitochondrial complexes I and V and adenosine triphosphate content, decreased expression of dynamin 1 protein, and increased expression of mitofusin 2 protein. Moreover, Si inhibited excessive hepatic apoptosis by regulating the B-cell lymphoma-2 (Bcl-2)/Bcl-2-associated-X-protein signaling pathway. Taken together, these results indicated that Si potentially mitigated PQ-induced hepatic oxidative insults by improving antioxidant capacity and mitochondrial function and inhibiting inflammation and cell apoptosis in weaned piglets.
... Also, another study found that silybin exerted antioxidant effects by regulating mitochondrial biogenesis in the liver of rats [33]. Mitochondria are counted as the factory of the cell, which generates ATP through oxidative phosphorylation relying on the respiratory chain, which is composed of five enzymatic complexes, including NADH dehydrogenase (complex I), succinate dehydrogenase (complex II), ubiquinol cytochrome c oxidoreductase (complex III), etc. [34]. A previous study showed that the PQ challenge causes protein oxidation in dopaminergic neurons by interfering with mitochondrial complex I activity [35]. ...

Oxidative stress is the major incentive for intestinal dysfunction in weaned piglets, which usually leads to growth retardation or even death. Silybin has caught extensive attention due to its antioxidant properties. Herein, we investigated the effect of dietary silybin supplementation on growth performance and determined its protective effect on paraquat (PQ)-induced intestinal oxidative damage and microflora dysbiosis in weaned piglets. In trial 1, a total of one hundred twenty healthy weaned piglets were randomly assigned into five treatments with six replicate pens per treatment and four piglets per pen, where they were fed basal diets supplemented with silybin at 0, 50, 100, 200, or 400 mg/kg for 42 days. In trial 2, a total of 24 piglets were randomly allocated to two dietary treatments with 12 replicates per treatment and 1 piglet per pen: a basal diet or adding 400 mg/kg silybin to a basal diet. One-half piglets in each treatment were given an intraperitoneal injection of paraquat (4 mg/kg of body weight) or sterile saline on day 18. All piglets were euthanized on day 21 for sample collection. The results showed that dietary supplementation with 400 mg/kg silybin resulted in a lower feed conversion ratio, diarrhea incidence, and greater antioxidant capacity in weaned piglets. Dietary silybin enhanced intestinal antioxidant capacity and mitochondrial function in oxidative stress piglets induced by PQ. Silybin inhibited mitochondria-associated endogenous apoptotic procedures and then improved the intestinal barrier function and morphology of PQ-challenged piglets. Moreover, silybin improved intestinal microbiota dysbiosis induced by the PQ challenge by enriching short-chain fatty-acid-producing bacteria, which augmented the production of acetate and propionate. Collectively, these findings indicated that dietary silybin supplementation linearly decreased feed conversion ratio and reduced diarrhea incidence in normal conditions, and effectively alleviated oxidative stress-induced mitochondrial dysfunction, intestinal damage, and microflora dysbiosis in weaned piglets.
... The basic function of mitochondria appears to be the production of ATP, which is necessary to support cellular exergonic activity. Aside from that, most vertebrate and invertebrate species have identified the whole collection of mitochondrial protein-coding genes revealed in humans, including COB (apocytochrome b), COX1-3 (cytochrome c oxidase subunits), ATP 6&8 (ATPase subunits), and NAD1-4,4 L,5,6 (NADH dehydrogenase subunits) ( Fig. 1) (Gissi, et al.2008;Bellance et al. 2009). Animal mitochondria may be defined as a small biologically inherited DNA, that encodes 37 genes, 13 key proteins of the oxidative phosphorylation (OXPHOS) system, and a further 24 genes are necessary for translation (Scheffler 2008a, b). ...




The peptides are known to have diverse therapeutic indications and help cure many disease conditions. The mitochondrial genome comprises a novel class of micro peptides with various biological effects. Human mitochondrial peptides are among those that have a favourable impact on medicinal applications. Mitochondrial-derived peptides are small therapeutic peptides transcribed through mitochondrial DNA open-reading frames. Humanin is a mitochondrial peptide inhibiting complex I activity and reducing oxidative stress. It has significant apoptosis activity; therefore, it acts as an anti-apoptotic agent in the testis, boosting its reproductive role. It primarily affects cardioprotective and neuroprotective pathways, which operate as Alzheimer’s disease treatment agents. SHLPs and humanin were derived from mitochondrially encoded 16s rRNA small open reading frames. This peptide enhances mitochondrial functions and the survival of cells in response to oxidative stress. SHLP (1–6) has distinct and overlapping functions with humanin which regulate cell viability. The anti-diabetic activity is present in SHLP 2 and 3, which are insulin-resistant. Likewise, SHLP-6 has an apoptotic effect on cancer cells while it protects the cells against mitochondrial-mediated apoptosis. The mitochondrial-derived peptides increase insulin sensitivity, stimulating glucose absorption through peripheral tissues while decreasing glucose synthesis in the liver. These peptides are helpful in the treatment of multiple diseases. Thus, in this review discusses the mitochondrial peptide-humanin and SHLP types and their therapeutic potential.
... For example, a deficiency in the ATP-synthase bsubunit was reported (López-Ríos et al., 2007). Often mitochondrial biogenesis is decreased, given by the decreased PGC1 expression, such as in lung (Bellance et al., 2009) and breast cancer (Watkins et al., 2004), consequently decreasing also mitochondrial transcription factor and mtDNA nucleoid structural protein TFAM. ...



Significance:
Mitochondrial (mt) reticulum network in the cell possesses amazing ultramorphology of parallel lamellar cristae, formed by the invaginated mitochondrial inner membrane (IMM). Its non-invaginated part, the inner boundary membrane (IBM) forms a cylindrical sandwich with the outer mitochondrial membrane (OMM). Crista membranes (CM) meet IBM at crista junctions (CJs) of mt cristae organizing system (MICOS) complexes connected to OMM SAM. Cristae dimensions, shape and CJs have characteristic patterns for different metabolic regimes, physiological, and pathological situations.
Recent advances:
Cristae-shaping proteins were characterized, namely rows of ATP-synthase dimers forming the crista lamella edges, MICOS subunits, OPA1 isoforms and MGM1 filaments, prohibitins and others. Detailed cristae ultramorphology changes were imaged by focused-ion beam/scanning electron microscopy. Dynamics of crista lamellae and mobile CJs were demonstrated by nanoscopy in living cells. With tBID-induced apoptosis a single entirely fused cristae reticulum was observed in a mitochondrial spheroid.
Critical issues:
The mobility and composition of MICOS, OPA1, and ATP-synthase dimeric rows regulated by posttranslational modifications might be exclusively responsible for cristae morphology changes, but ion fluxes across CM and resulting osmotic forces might be also involved. Inevitably, cristae ultramorphology should reflect also mitochondrial redox homeostasis, but details are unknown. Disordered cristae typically reflect higher superoxide formation.
Future directions:
To link redox homeostasis to cristae ultramorphology and define markers, recent progress will help in uncovering mechanisms involved in proton-coupled electron transfer via the respiratory chain and in regulation of cristae architecture, leading to structural determination of superoxide formation sites and cristae ultramorphology changes in diseases.
... In several studies, following oxidative DNA damage, mitochondria membrane potential collapse was realized in Cu(II) and Ni (II) complexes [53]. It should be noted that mitochondria have the main role in cellular energy metabolism by generating ATP via oxidative phosphorylation and also in the apoptosis process by translocating pro-apoptotic proteins from the mitochondrial intermembrane space to the cytosol [54]. In 2020. ...
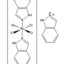



Two Cu(II) (C1) and Ni(II) (C2) complexes were designed through the one-pot reaction of pyridine-2,6-dicarboxylic acid and 2-aminobenzimidazole respectively with copper(II) nitrate hexahydrate and nickel(II) nitrate hexahydrate. Both complexes were characterized by single-crystal X-ray diffraction and the distorted octahedral geometry was recognized for them. The MTT assay indicated that the complexes have a significant antiproliferative effect on BEL-7404 cells. IC50 values confirmed that C1 (IC50 = 0.56 μM) is several times more potent than C2 (IC50 = 5.13 μM). The similar cellular uptake of the complexes in mentioned cells led to this proposal that the production of ROS with different values can be the main reason for different cytotoxicity of the complexes. In this study, C1 and C2 caused BEL-7404 cells arrest at the G2/M and S phases, respectively. The expression of p53, Bax up-regulation, and Bcl-2 down-regulation and also activation of procaspase-9, and 3 indicated that apoptosis through a caspase-dependent mitochondrion pathway is a remarkable pathway in BEL-7404 cells treated by C1 while mechanistic studies proved that C2 induce death of BEL-7404 cells through the activation of RAGE/PI3KC3/Beclin 1 autophagic cell signaling pathway, more specifically. The cytostatic effect of the complexes in the BEL-7404 3D spheroid model was depicted.
... Cellular ROS functions in signaling cascades as secondary messengers crucial for the general physiological performances known as differentiation and development [4,5]. However, excessive ROS production can also damage the body's biological substances such as DNA, carbohydrates, lipids, as well as proteins, leading to loss of cell integrity and subsequent cell pathology [6,7]. Clinical diseases such as inflammation, atherosclerosis, angiogenesis, aging, and cancer are influenced by abnormal ROS regulation [8]. ...





Reactive oxygen species (ROS) induce carcinogenesis by causing genetic mutations, activating oncogenes, and increasing oxidative stress, all of which affect cell proliferation, survival, and apoptosis. When compared to normal cells, cancer cells have higher levels of ROS, and they are responsible for the maintenance of the cancer phenotype; this unique feature in cancer cells may, therefore, be exploited for targeted therapy. Quercetin (QC), a plant-derived bioflavonoid, is known for its ROS scavenging properties and was recently discovered to have various antitumor properties in a variety of solid tumors. Adaptive stress responses may be induced by persistent ROS stress, allowing cancer cells to survive with high levels of ROS while maintaining cellular viability. However , large amounts of ROS make cancer cells extremely susceptible to quercetin, one of the most available dietary flavonoids. Because of the molecular and metabolic distinctions between malignant and normal cells, targeting ROS metabolism might help overcome medication resistance and achieve therapeutic selectivity while having little or no effect on normal cells. The powerful bioac-tivity and modulatory role of quercetin has prompted extensive research into the chemical, which has identified a number of pathways that potentially work together to prevent cancer, alongside, QC has a great number of evidences to use as a therapeutic agent in cancer stem cells. This current study has broadly demonstrated the function-mechanistic relationship of quercetin and how it Citation: Biswas, P.; Dey, D.; Biswas, P.K.; Rahaman, T.I.; Saha, S.; Parvez, A.; Khan, D.A.; Lily, N.J.; Saha, K.; Sohel, M.; et al. Publisher's Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. Copyright: © 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons At-tribution (CC BY) license (https://cre-ativecommons.org/licenses/by/4.0/). Int. J. Mol. Sci. 2022, 23, 11746 2 of 36 regulates ROS generation to kill cancer and cancer stem cells. Here, we have revealed the regulation and production of ROS in normal cells and cancer cells with a certain signaling mechanism. We demonstrated the specific molecular mechanisms of quercetin including MAPK/ERK1/2, p53, JAK/STAT and TRAIL, AMPKα1/ASK1/p38, RAGE/PI3K/AKT/mTOR axis, HMGB1 and NF-κB, Nrf2-induced signaling pathways and certain cell cycle arrest in cancer cell death, and how they regulate the specific cancer signaling pathways as long-searched cancer therapeutics.
... Biological systems are under a constant attack of reactive oxygen species. To defend such attacks, the body has a complex mechanism of regulating antioxidants, but when this system collapses, the biological system has to face oxidative stress which may lead to several diseased conditions such as cancer, aging, diabetes, inflammatory joint diseases and heart diseases [23,24]. This study aims to evaluate the medicinal potential of C. didymus extracts against cancer, diabetes, infectious bacteria and oxidative stress as well as the identification of major bioactive compounds present in these extracts. ...



Coronopus didymus (Brassicaceae) commonly known as lesser swine cress has been reported to be used for its pharmacological activities. This study aimed to evaluate the medicinal potential of C. didymus extracts against cancer, diabetes, infectious bacteria and oxidative stress and the identification of bioactive compounds present in these extracts. The effects of using different solvents for the extraction of C. didymus on the contents of major polyphenols and biological activities were investigated. Plant sample was shade dried, ground to a fine powder, and then soaked in pure acetone, ethanol and methanol. The highest contents of major polyphenols were found in methanol-based extract, i.e., chlorogenic acid, HB acid, kaempferol, ferulic acid, quercetin and benzoic acid with 305.02, 12.42, 11.5, 23.33, 975.7 and 428 mg/g of dry weight, respectively, followed by ethanol- and acetone-based extracts. The methanol-based extract also resulted in the highest antioxidant activities (56.76%), whereas the highest antiproliferative (76.36) and alpha glucosidase inhabitation (96.65) were demonstrated in ethanol-based extracts. No antibacterial property of C. didymus was observed against all the tested strains of bacteria. Further studies should be focused on the identification of specific bioactive compounds responsible for pharmacological activities.
... It has been shown that HIF-1α is not the only signal that induces the expression of glycolytic enzymes and therefore regulates glycolysis. Metabolites of glycolysis (e.g., pyruvate) and TAC (e.g., oxaloacetate, succinate, and fumarate) can also activate HIF-1α when they accumulate in the cell (because of violation of the ratios between glycolysis and oxidative phosphorylation) [25]. Among other things, pyruvate prevents the aerobic degradation of HIF-1α and enhances the expression of HIF-1α-activated genes, including erythropoietin, VEGF, glucose transporter-3 (GLUT-3), and aldolase A [26]. ...



The role of lactic acid (lactate) in cell metabolism has been significantly revised in recent decades. Initially, lactic acid was attributed to the role of a toxic end-product of metabolism, with its accumulation in the cell and extracellular space leading to acidosis, muscle pain, and other adverse effects. However, it has now become obvious that lactate is not only a universal fuel molecule and the main substrate for gluconeogenesis but also one of the most ancient metabolites, with a signaling function that has a wide range of regulatory activity. The Warburg effect, described 100 years ago (the intensification of glycolysis associated with high lactate production), which is characteristic of many malignant tumors, confirms the key role of lactate not only in physiological conditions but also in pathologies. The study of lactate’s role in the malignant transformation becomes more relevant in the light of the “atavistic theory of carcinogenesis,” which suggests that tumor cells return to a more primitive hereditary phenotype during microevolution. In this review, we attempt to summarize the accumulated knowledge about the functions of lactate in cell metabolism and its role in the process of carcinogenesis and to consider the possible evolutionary significance of the Warburg effect.
... During their oxidation, NADH and FADH 2 transfer their electrons, initiating the electron flux through the ETC, also known as respiratory chain ( Figure 3). This chain is made of 5 enzymatic complexes: NADH dehydrogenase (Complex I), succinate dehydrogenase (Complex II), ubiquinol cytochrome c oxidoreductase (Complex III), cytochrome c oxidase (Complex IV) and ATP synthase (Complex V) [42]. Electron transfer through ETC complexes is accompanied with proton pumping, from mitochondrial matrix toward IMS that generates electrical and chemical gradients. ...

Background
Systemic low-grade inflammation observed in diet-induced obesity has been associated with dysbiosis and disturbance of intestinal homeostasis. This latter relies on an efficient epithelial barrier and coordinated intestinal epithelial cell (IEC) renewal that are supported by their mitochondrial function. However, IEC mitochondrial function might be impaired by high fat diet (HFD) consumption, notably through gut-derived metabolite production and fatty acids, that may act as metabolic perturbators of IEC.
Scope of review
This review presents the current general knowledge on mitochondria, before focusing on IEC mitochondrial function and its role in the control of intestinal homeostasis, and featuring the known effects of nutrients and metabolites, originating from the diet or gut bacterial metabolism, on IEC mitochondrial function. It then summarizes the impact of HFD on mitochondrial function in IEC of both small intestine and colon and discusses the possible link between mitochondrial dysfunction and altered intestinal homeostasis in diet-induced obesity.
Major conclusions
HFD consumption provokes a metabolic shift toward fatty acid β-oxidation in the small intestine epithelial cells and impairs colonocyte mitochondrial function, possibly through downstream consequences of excessive fatty acid β-oxidation and/or the presence of deleterious metabolites produced by the gut microbiota. Decreased levels of ATP and concomitant O2 leaks into the intestinal lumen could explain the alterations of intestinal epithelium dynamics, barrier disruption and dysbiosis that contribute to the loss of epithelial homeostasis in diet-induced obesity. However, the effect of HFD on IEC mitochondrial function in the small intestine remains unknown and the precise mechanisms by which HFD induces mitochondrial dysfunction in the colon have not been elucidated so far.