Fig 24 - uploaded by C. H. J. M. Fransen
Content may be subject to copyright.

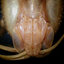
Map of the underwater cave off N coast of Lavernaka Island, Kornati Archipelago, Middle Dalmatia, Adriatic, indicating sites where specimens of Salmoneus sketi where seen (1-3) and caught (1,3). Drawing by B. Sket. Colour.-Transparent except for orange-red hepatopancreas, eyes and eggs (B. Sket personal communication; colour slides). Description of ovigerous female paratype as far as different from holotype.-Rostrum elongate, about one third of pocl., reaching half of third seg-ment of antennular peduncle, not ca-rinate dorsally; one ventral tooth pre-sent (figs. 25, 26). Lateral margins of rostrum concave in dorsal aspect (fig. 27). Lateral eaves partly concealing eyes in dorsal view (fig. 27). Orbital teeth sharp. Antennular peduncle reaching beyond tip of rostrum. Stylocerite not over-reaching second antennular segment. Mouthparts not studied. Chela of left first pereiopod (fig. 28) with six small triangular teeth in distal half of movable finger. Fixed finger with five small triangular teeth in distal half and eleven small teeth in proximal half. Habitat.-The locality where the specimens were caught is a small, vertical, marine cave with the entrance at 7 m and the bottom at 20 m depth approximately (fig. 24). Walls are partly covered by dripstone formations as well as by fingerlike formations of bryozoan origin, in some sheltered places very long tubes of Polychaeta (Filogranula cf. annulata
Source publication






A new species of Alpheidae, Salmoneus sketi, is described and figured. Two specimens of the species were found in a submarine cave in the Adriatic.
Similar publications








Despite their role in marine systems, Sergestidae remain one of the most poorly understood families amongst planktonic shrimps with regard to phylogeny. Recent morphological and phylogenetic revisions of a number of sergestid genera have disentangled classificatory problems and emphasized the importance of reproductive structures for the taxonomy a...








The fossil shrimp Araripenaeus timidus n. gen. n. sp. is the first fossil Penaeoidea from Brazil. Here, we describe, illustrate and compare it with Paleomattea deliciosa, a fossil Sergestoidea (Dendrobranchiata) previously described from the same region. The material of the early Cretaceous (Albian) was collected in the town of Jardim, south of the...









The mainland of Sulawesi is well known for its high diversity of freshwater shrimps. However, many small islands surrounding Sulawesi have been neglected and have not been unexplored. Here, we report on the freshwater shrimps of the Banggai Archipelago, eastern Sulawesi, based on field collections between 2017 and 2019 on four islands, Peleng, Labo...







New records are presented for the caridean shrimp family Processidae in Indo-West and East Pacific waters, based mainly on recently collected material. These records highlight that many processid species may be relatively widespread, although the distribution of some species remains imperfectly known. The colour pattern of several species is here d...






The content of As, Cd, Cr, Cu, Fe, Mn, Ni, Pb, and Zn as the whole-body burden and the content of Cd and Pb in muscle tissue of Parapenaeus longirostris, were evaluated in shrimp collected from the Pagasitikos Gulf, Greece. The whole-body burden of Fe was 102 ± 39, Mn 71 ± 23, As 64 ± 33, Zn 62 ± 8, Cu 34 ± 16, Ni 4.6 ± 2.9, Cr 1.01 ± 0.39, Cd 0.93...
Citations
... However, the actual species diversity of the genus appears to be far beyond what is currently known, considering the presence of numerous morphologically nearly identical species, the rapidly growing number of described species in the genus, and the fact that about two-thirds of all the species have been described in the past two decades (e.g., Anker 2010; Komai and Anker 2012;Anker and Lazarus 2015;Anker 2019b;Anker and Ashrafi 2019;Anker et al. 2020;Marin 2021;Anker 2022;Komai 2022). The reason for this hidden diversity lies in these shrimps' small size and cryptic lifestyles, ranging from dwelling in small spaces under large boulders or in coral rubble crevices (Anker and Marin 2006;Anker 2011Anker 2019aAnker 2020Ashrafi et al. 2020) to living in burrows of various fossorial animals (e.g., De Grave 2004;Komai 2009;Anker et al. 2014;Komai et al. 2021;d'Acoz et al. 2022), or in marine caves (Fransen 1991;Komai et al. 2015). ...
... Etymology: The species is named after Dr. Charles H.J.M. Fransen (Naturalis Biodiversity Center, Leiden) for his major contribution to taxonomy of caridean shrimps, including description of a peculiar cave species of Salmoneus (Fransen 1991). ...



... Previous research reported large individuals of S. carvachoi brooding embryos underneath the pleon and exhibiting appendices masculinae on the endopods of the second pleopods (Oliveira et al. 2015, Vera-Caripe et al. 2015. The same condition has been observed before in other congeneric species (Christoffersen 1982, Carvacho 1989, Holthuis 1990, Fransen 1991, Anker and Marin 2006, Anker 2007, 2010, 2011a. Furthermore, ovigerous shrimps of S. carvachoi (bearing appendices masculinae) are commonly found sharing the same refuge (Oliveira et al. 2015, Guéron et al. 2022). ...
... For instance, the appendix masculina (AM) is absent in all individuals in the alpheid genera Automate De Man, 1888 and Synalpheus Spence Bate, 1888 species (Banner and Banner 1973, Felder 1982, Dardeau 1984, Carvacho 1989, Tóth and Bauer 2007. On the other hand, in Salmoneus and Yagerocaris Kensley, 1988, the AM is present in all individuals regardless of the presence or absence of eggs brooded in their pleon (Christoffersen 1982, Carvacho 1989, Holthuis 1990, Fransen 1991, Anker and Marin 2006, Anker 2007, 2010, 2011a, b, Olivera et al. 2015, Vera-Caripe et al. 2015. Carvacho (1989) stated that if the presence of the AM in Salmoneus was an expression of protandry, then, a gradual regression of this structure should be expected after sexual inversion. ...





In caridean shrimps, sexual systems vary from gonochorism to various forms of hermaphroditism. The shrimp Salmoneus carvachoi has been reported to exhibit both male (appendix masculina) and female (brooding embryos) anatomical characteristics simultaneously. Our aim was to examine the sexual system of S. carvachoi and to test the hypothesis of protandric simultaneous hermaphroditism (PSH) in this species. We analysed internal and external features using microscopy and histology techniques. Our analyses support the hypothesis of PSH in S. carvachoi and enabled the identification of male-phase and hermaphroditic individuals. All individuals exhibit a well-developed appendix masculina on the endopods of the second pleopods; however, small male-phase individuals possess only male gonopores, whereas larger hermaphroditic individuals possess both female and male gonopores. Hermaphrodites exhibit both male and female germinative cells in various stages of development, as well as an androgenic gland. Our study describes a novel sexual system for alpheids and provides a detailed description of anatomical traits in S. carvachoi. Studies that describe sexual systems in Alpheidae may shed light on the evolutionary history of gender expression in carideans and how environmental conditions favour sex allocation in decapods and other invertebrates.
... Representatives of Salmoneus typically have five pleurobranchs, the presence of arthrobranch at the base of the third maxilliped, the absence of a podobranch, sac-shaped epipods on the first and second maxillipeds, epipods on the third maxillipeds to the fourth pereiopods, and setobranchs on all pereiopods [9,10]. However, modification in arthrobranch, such as a notably reduced size or hypertrophy, can be observed among some members of the genus, albeit quite infrequently [18,19]. ...
... However, assessing the size of the arthrobranch may not always be straightforward, as several previous studies have not given careful attention to gill size. The majority of species possess well-developed arthrobranchs [6,17,23], and instances of hypertrophy or remarkable reduction are uncommon [18,19]. Salmoneus sketi Fransen, 1991, and S. chelocrassus sp. ...
... Clear distinctive characters include a robust rostrum, possession of both major and subminor chelipeds in S. sketi, in contrast to S. chelocrassus sp. nov., possessing major and minor chelipeds, and a distinctive structure and arrangement of teeth along the major cheliped's cutting edge [18]. ...

During a faunal investigation using SCUBA in Taiwan in 2014, a single ovigerous specimen
of an alpheid shrimp belonging to the genus Salmoneus Holthuis, 1955, was collected. Notably, the
specimen stood out due to its extremely robust major cheliped compared to other species within the
genus. Detailed examination of the specimen unveiled its morphological distinctiveness from all
known Salmoneus species, confirming its novelty to science. Apart from the remarkably inflated major
cheliped devoid of depressions along the ventral margin, the characteristics of S. chelocrassus sp. nov.,
include the markedly reduced arthrobranch on the third maxilliped and the absence of microserrulate
setae on the propodus of the fifth pereiopod.
... At least 20 species of Salmoneus are infaunal shrimps confined to near-shore sandflats and mudflats, including estuaries and mangroves (Anker 2003a(Anker , 2003b(Anker , 2007Anker & Lazarus 2015;Marin 2021), where they often associate with various larger burrowing animals, such as alpheid shrimps of the genus Alpheus Fabricius, 1798 (De Grave 2004;Anker 2007;Anker & Marin 2006;Anker & Lazarus 2015;Anker & De Grave 2016), callianassid ghost shrimps (Dworschak et al. 2000;Anker 2003bAnker , 2011aAnker , 2019bKomai 2009;Anker & Ashrafi 2019;Anker et al. 2020;d'Udekem d'Acoz et al. 2022), upogebiid mud-shrimps (Marin 2021), and stomatopods (Anker 2019a). In addition, a few species of Salmoneus are known from marine caves (Fransen 1991;Komai et al. 2015). The rapidly growing number of species in Salmoneus over the past few years suggests that the presently known diversity of the genus is still very far from the real one. ...
During a survey of the mangrove infauna on the Iranian side of the Persian Gulf and the Gulf of Oman several specimens of a new alpheid shrimp, Salmoneus shojaei sp. nov., were collected around pneumatophores of mangrove trees, mostly in association with the larger burrowing snapping shrimps of the genus Alpheus Fabricius, 1798. The new species appears to be a member of the S. gracilipes species group and is morphologically closest to S. colinorum De Grave, 2004 and S. alpheophilus Anker & Marin, 2006. However, a unique combination of morphological characters, such as the carapace without rostral carina, the unarmed ischium of the major cheliped, the armed ischia of the minor cheliped and second pereiopod, the very slender dactyli of the fourth and third pereiopods, and the posterior margin of the telson with a deep U-shaped notch, distinguishes the new species from all other members of the S. gracilipes group. In addition, S. shojaei sp. nov. presents a diagnostic, albeit very faint, banding of the pleon, which separates it from most other species of the S. gracilipes group with known colour patterns. A DNA barcode (a partial fragment of the mitochondrial gene, CO1), as well as partial fragments of the mitochondrial 16S rRNA and the nuclear H3 genes, are provided to genetically characterise the new taxon.
... In semi-submerged marine caves of Montenegro, Mačić et al. [31] found five species. Two species new to science were described from Mediterranean marine caves: Odontozona addaia, from Menorca (Balearic Islands, Spain) [32], and Salmoneus sketi, from Lavernaka Island (Croatia) [33]. ...
... Three are long known to live exclusively in the Mediterranean Sea (see Table S21 by C. Froglia in [55]): Hippolyte holthuisi, which has recently been recognized as the Mediterranean vicariant of the Atlantic congeneric H. varians Leach, 1814 [56], Maja squinado and Periclimenes amethysteus. Odontozona addaia and Salmoneus sketi are hitherto known only from the Mediterranean Sea [32][33][34]. Finally, Caridion sp. was originally identified with the boreal species C. steveni Lebour, 1930 by Ledoyer [57], who found it in a cave at Villefranche-sur-Mer (France), but considered as a distinct, still undescribed species by subsequent authors [50,58]. Lebour, 1930 by Ledoyer [57], who found it in a cave at Villefranche-sur-Mer (France), considered as a distinct, still undescribed species by subsequent authors [50,58]. ...
... Morphological and behavioural adaptations to cave life typical of stygobiotic species can be observed in caridean and stenopodid shrimps as well as in galatheid and brachyuran crabs found in marine caves [103]. Two species have not been-at least yet-reported from habitats other than marine caves and thus could perhaps be considered as 'stygobionts' (i.e., cave-exclusive): the stenopodid Odontozona addaia, from the Balearic Sea and the Gulf of Lion [34], and the alpheid Salmoneus sketi, from the Adriatic Sea [33]. Future research in cryptic and deep-sea habitats might substantiate or refute their presumed stygobiosis. ...


Decapod crustaceans are important components of the fauna of marine caves worldwide, yet information on their ecology is still scarce. Mediterranean marine caves are perhaps the best known of the world and may offer paradigms to the students of marine cave decapods from other geographic regions. This review summarizes and updates the existing knowledge about the decapod fauna of Mediterranean marine caves on the basis of a dataset of 76 species from 133 caves in 13 Mediterranean countries. Most species were found occasionally, while 15 species were comparatively frequent (found in at least seven caves). They comprise cryptobiotic and bathyphilic species that only secondarily colonize caves (secondary stygobiosis). Little is known about the population biology of cave decapods, and quantitative data are virtually lacking. The knowledge on Mediterranean marine cave decapods is far from being complete. Future research should focus on filling regional gaps and on the decapod ecological role: getting out at night to feed and resting in caves during daytime, decapods may import organic matter to the cave ecosystem. Some decapod species occurring in caves are protected by law. Ecological interest and the need for conservation initiatives combine to claim for intensifying research on the decapod fauna of the Mediterranean Sea caves.
... In semi-submerged marine caves of Montenegro, Mačić et al. [31] found five species. Two species new to science were described from Mediterranean marine caves: Odontozona addaia, from Menorca (Balearic Islands, Spain) [32], and Salmoneus sketi, from Lavernaka Island (Croatia) [33]. ...
... Three are long known to live exclusively in the Mediterranean Sea (see Table S21 by C. Froglia in [55]): Hippolyte holthuisi, which has recently been recognized as the Mediterranean vicariant of the Atlantic congeneric H. varians Leach, 1814 [56], Maja squinado and Periclimenes amethysteus. Odontozona addaia and Salmoneus sketi are hitherto known only from the Mediterranean Sea [32][33][34]. Finally, Caridion sp. was originally identified with the boreal species C. steveni Lebour, 1930 by Ledoyer [57], who found it in a cave at Villefranche-sur-Mer (France), but considered as a distinct, still undescribed species by subsequent authors [50,58]. Lebour, 1930 by Ledoyer [57], who found it in a cave at Villefranche-sur-Mer (France), considered as a distinct, still undescribed species by subsequent authors [50,58]. ...
... Morphological and behavioural adaptations to cave life typical of stygobiotic species can be observed in caridean and stenopodid shrimps as well as in galatheid and brachyuran crabs found in marine caves [103]. Two species have not been-at least yet-reported from habitats other than marine caves and thus could perhaps be considered as 'stygobionts' (i.e., cave-exclusive): the stenopodid Odontozona addaia, from the Balearic Sea and the Gulf of Lion [34], and the alpheid Salmoneus sketi, from the Adriatic Sea [33]. Future research in cryptic and deep-sea habitats might substantiate or refute their presumed stygobiosis. ...
Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review
... Salmoneus Holthuis 1955 is a genus of small-sized alpheid shrimps distributed throughout the tropical, subtropical and warm-temperate seas of the world, currently comprising 58 species (e.g., De Grave & Fransen 2011;Anker 2010Anker , 2011Anker , 2013Anker , 2019aAnker , b, 2020Ďuriš & Horká 2016). Prior to the present study, Salmoneus had only been recorded in the temperate North-East Atlantic from the Mediterranean Sea (Holthuis & Gottlieb 1958;Fransen 1991;Dworschak et al. 2000;Grippa 2004), but not from the European coast of the Atlantic Ocean. ...
... The taxonomy of S. serratidigitus itself is not fully resolved (Anker et al. 2015;Anker 2019b). (Fransen 1991). At least four of several morphological differences between the holotype and the paratype of S. sketi (see Fransen 1991: figs. ...

The rare symbiotic alpheid shrimp Salmoneus erasimorum Dworschak, Abed-Navandi & Anker, 2000 was previously known from a single specimen collected with a suction pump on the Croatian coast in the Adriatic Sea, together with its host, the ghost shrimp, Gilvossius tyrrhenus (Petagna, 1792). A second record of S. erasimorum is presented here, with a diagnosis and the first colour photographs, based on a single specimen collected in northern Brittany, France, also with a suction pump, but without its host. This is also the first record of the species on the European coast of the Atlantic Ocean. An annotated list and a key to the species of Salmoneus currently known from the eastern Atlantic and the Mediterranean Sea are provided.
... From the very first stages of marine cave research, it became evident that this peculiar habitat harbours several previously undescribed species (Sarà 1958, Vacelet & Lévi 1958, Vacelet 1959, Sarà & Siribelli 1960, Rützler & Sarà 1962. Until today, studies in Mediterranean marine and anchialine caves are continuously bringing to light new species from various taxonomic groups: Porifera (Vacelet & Boury-Esnault 1982, Pulitzer-Finali 1983, Pansini 1984, Bibiloni 1993, Corriero et al. 1996, 1997a, Bavestrello et al. 1997, Muricy et al. 1998, Pansini & Pesce 1998, Vacelet & Pérez 1998, Vacelet et al. 2000, 2007, Manconi et al. 2006, Pérez et al. 2011, Pisera & Vacelet 2011, Reveillaud et al. 2012, Melis et al. 2016, Lage et al. 2018, Priapulida (Todaro & Shirley 2003), Gastrotricha (Fregni et al. 1998), Copepoda , Carola & Razouls 1996, Jaume & Boxshall 1996, Jaume 1997, Jaume et al. 1999, Krsinic 2005, Decapoda (Pretus 1990, Fransen 1991, Mysida (Alcaraz et al. 1986, Wittmann 2004), Thermosbaenacea (Wagner & Chevaldonné 2020), Polychaeta (Zibrowius 1968, Fassari & Mòllica 1991, Gastropoda (Warén et al. 1997, Palazzi & Villari 2001, Crocetta et al. 2020), Bivalvia (La Perna 1998, Tardigrada (Villora-Moreno 1996), Bryozoa (Hayward 1974, Silén & Harmelin 1976, Harmelin et al. 2007, Rosso et al. 2020a), Brachiopoda (Logan & Zibrowius 1994), Chaetognatha (Casanova 1986) and Pisces (Kovačić 1999). ...
... In addition, many new copepods, decapods and a thermosbaenacean species, probably all cave-exclusive, were found from marine and anchialine caves of the Adriatic Sea, the Balearic Islands and south France (e.g. Pretus 1990, Fransen 1991, Jaume & Boxshall 1996, Jaume 1997, Jaume et al. 1999, Wagner & Chevaldonné 2020. The molecular study of mysids in several marine caves from different Mediterranean regions by Rastorgueff et al. (2014) revealed at least four cryptic Hemimysis taxa still undescribed. ...
Marine caves are biodiversity reservoirs and refuge habitats, harbouring rare species and living fossils. The Mediterranean Sea hosts more than 3000 caves, which are among the most studied in the world. This review aims to synthesize and update knowledge of Mediterranean marine caves. Their biota includes few obligate cave-dwelling organisms, but many cryptobiotic or crevicular (crevice-dwelling) and bathyphilic (preferring deep-water) species that secondarily colonize caves. A total of 2369 taxa have been reported from 404 caves in 15 countries, with several species new to science described in recent decades. Dramatic environmental gradients generate a zonation of the biota, with up to six faunal zones and two main biocoenoses. Biotic cover and biomass are strongly reduced inside caves, due to hydrological confinement and trophic depletion. The food web is based on suspension-feeders, but motile carnivores play a role in the importation of organic matter from outside. Lack of primary production, faunal affinities and microbial metabolism make marine caves readily accessible models of deep ocean ecosystems. Future research should focus on filling regional (e.g. south-eastern Mediterranean) and thematic (e.g. microbes, meiofauna, macroinfauna) gaps in fundamental knowledge, and on management measures. Marine caves have low ecological resilience and harbour many species of conservation interest, but are threatened by seawater warming, local human impacts and non-indigenous species.
... These features, however, are also present in some other species of Salmoneus (e.g., Anker & Marin 2006;. On the other side, in the general shape of the major cheliped, especially the cup-shaped, non-elongate carpus, S. singularis n. sp. is closer to almost all other species currently placed in Salmoneus, except for the cavernicolous S. sketi Fransen, 1991 andS. antricola Komai, Yamada &Yunokawa, 2015, in which the carpus is vase-shaped or long and cylindrical, as in Triacanthoneus (Fransen 1991;. ...
... On the other side, in the general shape of the major cheliped, especially the cup-shaped, non-elongate carpus, S. singularis n. sp. is closer to almost all other species currently placed in Salmoneus, except for the cavernicolous S. sketi Fransen, 1991 andS. antricola Komai, Yamada &Yunokawa, 2015, in which the carpus is vase-shaped or long and cylindrical, as in Triacanthoneus (Fransen 1991;. It is important to note here that the development of the notch on the posterior margin of the telson is variable within both genera (Anker 2010b(Anker , 2020aAlvarez et al. 2012Alvarez et al. , 2014; however, most species of Triacanthoneus do not have one, whereas most species of Salmoneus present a fairly deep notch, as is the case of S. singularis n. sp. ...

A new alpheid shrimp species, provisionally assigned to the genus Salmoneus Holthuis, 1955, is described and illustrated on the basis of three specimens extracted from intertidal soft sediments on seagrass beds at Iriomote Island, southern Ryukyu Islands, Japan. Salmoneus singularis n. sp. is highly distinctive within the genus in having a pair of postorbital spines on the carapace, otherwise showing some affinities to the species of the S. gracilipes Miya, 1972 group. The possession of paired spines on the dorsolateral surface of the carapace is shared with several members of Triacanthoneus Anker, 2010, which is undoubtedly closely related to Salmoneus. However, the new species cannot be placed in Triacanthoneus because of the absence of a mid-dorsal spine on the carapace.
... The genus is near-cosmopolitan in distribution, occurring in tropical to temperate latitudes, with species known from the Mediterranean Sea, West Africa, islands of the central Atlantic, Brazil, the wider Caribbean-Florida-Gulf of Mexico area, tropical eastern Pacific, as well as across the Indo-West Pacific from the Red Sea and South Africa through to Japan, Hawaii and French Polynesia (see references above). Ecologically, a wide number of niches are occupied, with cave dwellers (e.g. S. sketi Fransen, 1991;S. antricola Komai, Yamada & Yunokawa, 2015) and inhabitants of mangroves and mudflats (e.g. S. seticheles Anker, 2003;S. ...


The alpheid shrimp Salmoneus tafaongae Banner & Banner, 1966 was described based on a fragmentary holotype from Samoa, which was subsequently lost in the devastating fire of the Hawaii Marine Laboratory in 1961. The species is here fully redescribed based on new material from French Polynesia, Australia (Great Barrier Reef) and Solomon Islands, and compared to closely related species. LSID urn:lsid:zoobank.org:pub:441C3DC5-8B7B-40BE-89D4-577EFB939FD2