FIG 2 - uploaded by Else Vellinga
Content may be subject to copyright.

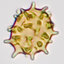
Lepiota kauffmanii. A. Spores. B. Cheilocystidia. C. Pileus covering elements. All from the holotype (Zeller 7806). Bar 5 10 mm.
Source publication






Eight Lepiota species with a hymeniform pileus covering that are known in California are presented and discussed. Lepiota phaeoderma is described as new; this species is characterized by a dark pileus surface, which splits open into small patches around the umbo, and the absence of an annulus. Lepiota neophana, a species with an annulus and a close...
Similar publications







In this report we describe the three species in Lepiota sect. Lepiota occurring in tropical China. Lepiota attenuata is a new species and is characterized by a pileus with brownish yellow squamules and radially sulcate striate margin, penguin-shaped spores that are distinctively narrowed toward the apex and inflated submoniliform or catenulate elem...
Citations
... The phylogenetic tree in Figure 1 shows the relationship between the lepiotaceous species included in this study. All sequences of the specimens from Hungary clustered together with a L. cystophora sequence (GQ141550) originated from a specimen collected in Italy and published as Cystolepiota cystophora (Vellinga 2010;Osmundson et al. 2013) in a strongly supported clade (MLBS = 99%). Lepiota cystophora formed a strongly supported sister clade (MLBS = 98%) in our analysis represented by a sequence of Lepiota cystophoroides Joss. ...
... The known sequences of the morphologically identified Cystolepiota species did not form a monophyletic group across any phylogeny published to date (e.g. Vellinga 2003bVellinga , 2010Qu et al. 2023). One of the key problems to understand the delimitation of Cystolepiota, is the lack of any sequence data of C. constricta, the type species of the genus. ...


... Fayod even erected the genus Fusispora Fayod to accommodate L. sistrata and considered A. seminudus as a member of another new genus, Cystoderma [25]. In contrast, Vellinga reduced L. sororia Huijsman and L. rufipes Morgan to synonymy with C. seminuda [21,36]. ...
... Compared with C. seminuda, L. sororia and L. rufipes, which have been reduced to synonyms of C. seminuda by Vellinga in 1987 and 2010 respectively, have larger basidiomata and longer and narrower basidiospores [21,33,36,59]. Furthermore, L. sororia forms larger basidiomata [33] and both the pileus and stipe of L. rufipes were described as smooth and glabrous [59], although inferred that it may be the result of rain washing [36]. ...
... Compared with C. seminuda, L. sororia and L. rufipes, which have been reduced to synonyms of C. seminuda by Vellinga in 1987 and 2010 respectively, have larger basidiomata and longer and narrower basidiospores [21,33,36,59]. Furthermore, L. sororia forms larger basidiomata [33] and both the pileus and stipe of L. rufipes were described as smooth and glabrous [59], although inferred that it may be the result of rain washing [36]. Accounting for the difference described above, we temporarily remove L. sororia and L. rufipes from synonyms of C. seminuda until specimens exactly matching the description of the protologue of the two species are found and sequenced. ...



Species of Cystolepiota are known as diminutive lepiotaceous fungi with a worldwide distribution. Previous studies revealed that Cystolepiota is not monophyletic and preliminary DNA sequence data from recent collections suggested that several new species exist. Based on multi-locus DNA sequence data (the nuc rDNA internal transcribed spacer region ITS1-5.8S-ITS2, ITS; the D1–D2 domains of nuc 28S rDNA, LSU; the most variable region of the second-largest subunit of RNA polymerase II, rpb2 and a portion of the translation–elongation factor 1-α. tef1), C. sect. Pulverolepiota forms a distinct clade separating from Cystolepiota. Therefore, the genus Pulverolepiota was resurrected and two combinations, P. oliveirae and P. petasiformis were proposed. With the integration of morphological characteristics, multi-locus phylogeny, and information on geography and habitat, two new species, viz. C. pseudoseminuda and C. pyramidosquamulosa, are described and C. seminuda was revealed to be a species complex containing at least three species, viz. C. seminuda, C. pseudoseminuda, and Melanophyllum eryei. In addition, C. seminuda was re-circumscribed and neo-typified based on recent collections.
... Alignment and phylogenetic analysis:-Sequences obtained from the new species were compared with sequences in GenBank and UNITE (https://unite.ut.ee/) databases. Sequences were retrieved from GenBank and UNITE for phylogenetic analysis based on the Basic Local Alignment Search Tool (BLAST) results with the highest similarity (maximum identities greater than 90%) to the new species and the outcomes of previous phylogenetic studies involving Lepiota sequences (Johnson 1999, Vellinga 2001a, 2001c, 2003, 2006a, 2006b, Hughes et al. 2009, Liang et al. 2010, Vellinga 2010, Birkebak et al. 2011, Sysouphanthong et al. 2011, Kropp et al. 2012, Sysouphanthong et al. 2012, Ge & Smith 2013, Lebel & Vellinga 2013, Caballero et al. 2015, Vidal et al. 2015, Liang et al. 2018, Hou & Ge 2020and Long et al. 2020. In addition, the sequences of morphologically similar taxa to the new species were also included in the data matrix. ...

A new species of Lepiota from Turkey, Lepiota cremea sp. nov., is proposed based on morphological characters and phylogenetic analyses of the nuclear rDNA internal transcribed spacer (ITS1-5.8S-ITS2 = ITS) region. The morphological features together with molecular data confirmed both the novelty of this species and its infrageneric placement within Lepiota sect. Lepiota. Lepiota cremea, so far only found in Pinus nigra subsp. pallasiana forest, is characterized by the pale beige basidiomata, subcylindrical to fusiform basidiospores, long pileus covering elements with often nodulose or horn-like bumps. A comprehensive description, illustrations and comparisons with morphologically similar and related taxa are provided.
... The only differences of our species from the latter are a smooth, non-wrinkled pileus, persistent and funnel-shaped annulus on the stem, which is flimsy and evanescent in L. neophana, and the presence of 2-spored basidia. According to E.C. Vellinga (Vellinga 2010 likely represents a species complex and may combine several species that differ phylogenetically and in their geographic distribution, and our species is very close to this complex (see phylogenetic tree below). The ITS sequences from L. neo phana show a clear genetic distance from L. sayanensis that reaches 4 %. ...



... The only differences of our species from the latter are a smooth, non-wrinkled pileus, persistent and funnel-shaped annulus on the stem, which is flimsy and evanescent in L. neophana, and the presence of 2-spored basidia. According to E.C. Vellinga (Vellinga 2010 likely represents a species complex and may combine several species that differ phylogenetically and in their geographic distribution, and our species is very close to this complex (see phylogenetic tree below). The ITS sequences from L. neo phana show a clear genetic distance from L. sayanensis that reaches 4 %. ...

Novel species of fungi described in this study include those from various countries as follows: Antartica, Cladosporium austrolitorale from coastal sea sand. Australia, Austroboletus yourkae on soil, Crepidotus innuopurpureus on dead wood, Curvularia stenotaphri from roots and leaves of Stenotaphrum secundatum and Thecaphora stajsicii from capsules of Oxalis radicosa. Belgium, Paraxerochrysium coryli (incl. Paraxerochrysium gen. nov.) from Corylus avellana. Brazil, Calvatia nordestina on soil, Didymella tabebuiicola from leaf spots on Tabebuia aurea, Fusarium subflagellisporum from hypertrophied floral and vegetative branches of Mangifera indica and Microdochium maculosum from living leaves of Digitaria insularis. Canada, Cuphophyllus bondii from a grassland. Croatia, Mollisia inferiseptata from a rotten Laurus nobilis trunk. Cyprus, Amanita exilis on calcareous soil. Czech Republic, Cytospora hippophaicola from wood of symptomatic Vaccinium corymbosum. Denmark, Lasiosphaeria deviata on pieces of wood and herbaceous debris. Dominican Republic, Calocybella goethei among grass on a lawn. France (Corsica), Inocybe corsica on wet ground. France (French Guiana), Trechispora patawaensis on decayed branch of unknown angiosperm tree and Trechispora subregularis on decayed log of unknown angiosperm tree. Germany, Paramicrothecium sambuci (incl. Paramicrothecium gen. nov.) on dead stems of Sambucus nigra. India, Aureobasidium microtermitis from the gut of a Microtermes sp. termite, Laccaria diospyricola on soil and Phylloporia tamilnadensis on branches of Catunaregam spinosa. Iran, Pythium serotinoosporum from soil under Prunus dulcis. Italy, Pluteus brunneovenosus on twigs of broadleaved trees on the ground. Japan, Heterophoma rehmanniae on leaves of Rehmannia glutinosa f. hueichingensis. Kazakhstan, Murispora kazachstanica from healthy roots of Triticum aestivum. Namibia, Caespitomonium euphorbiae (incl. Caespitomonium gen. nov.) from stems of an Euphorbia sp. Netherlands, Alfaria junci, Myrmecridium junci, Myrmecridium juncicola, Myrmecridium juncigenum, Ophioceras junci, Paradinemasporium junci (incl. Paradinemasporium gen. nov.), Phialoseptomonium junci, Sporidesmiella juncicola, Xenopyricularia junci and Zaanenomyces quadripartis (incl. Zaanenomyces gen. nov.), from dead culms of Juncus effusus, Cylindromonium everniae and Rhodoveronaea everniae from Evernia prunastri, Cyphellophora sambuci and Myrmecridium sambuci from Sambucus nigra, Kiflimonium junci, Sarocladium junci, Zaanenomyces moderatricis-academiae and Zaanenomyces versatilis from dead culms of Juncus inflexus, Microcera physciae from Physcia tenella, Myrmecridium dactylidis from dead culms of Dactylis glomerata, Neochalara spiraeae and Sporidesmium spiraeae from leaves of Spiraea japonica, Neofabraea salicina from Salix sp., Paradissoconium narthecii (incl. Paradissoconium gen. nov.) from dead leaves of Narthecium ossifragum, Polyscytalum vaccinii from Vaccinium myrtillus, Pseudosoloacrosporiella cryptomeriae (incl. Pseudosoloacrosporiella gen. nov.) from leaves of Cryptomeria japonica, Ramularia pararhabdospora from Plantago lanceolata, Sporidesmiella pini from needles of Pinus sylvestris and Xenoacrodontium juglandis (incl. Xenoacrodontium gen. nov. and Xenoacrodontiaceae fam. nov.) from Juglans regia. New Zealand, Cryptometrion metrosideri from twigs of Metrosideros sp., Coccomyces pycnophyllocladi from dead leaves of Phyllocladus alpinus, Hypoderma aliforme from fallen leaves Fuscopora solandri and Hypoderma subiculatum from dead leaves Phormium tenax. Norway, Neodevriesia kalakoutskii from permafrost and Variabilispora viridis from driftwood of Picea abies. Portugal, Entomortierella hereditatis from a biofilm covering a deteriorated limestone wall. Russia, Colpoma junipericola from needles of Juniperus sabina, Entoloma cinnamomeum on soil in grasslands, Entoloma verae on soil in grasslands, Hyphodermella pallidostraminea on a dry dead branch of Actinidia sp., Lepiota sayanensis on litter in a mixed forest, Papiliotrema horticola from Malus communis, Paramacroventuria ribis (incl. Paramacroventuria gen. nov.) from leaves of Ribes aureum and Paramyrothecium lathyri from leaves of Lathyrus tuberosus. South Africa, Harzia combreti from leaf litter of Combretum collinum ssp. sulvense, Penicillium xyleborini from Xyleborinus saxesenii, Phaeoisaria dalbergiae from bark of Dalbergia armata, Protocreopsis euphorbiae from leaf litter of Euphorbia ingens and Roigiella syzygii from twigs of Syzygium chordatum. Spain, Genea zamorana on sandy soil, Gymnopus nigrescens on Scleropodium touretii, Hesperomyces parexochomi on Parexochomus quadriplagiatus, Paraphoma variabilis from dung, Phaeococcomyces kinklidomatophilus from a blackened metal railing of an industrial warehouse and Tuber suaveolens in soil under Quercus faginea. Svalbard and Jan Mayen, Inocybe nivea associated with Salix polaris. Thailand, Biscogniauxia whalleyi on corticated wood. UK, Parasitella quercicola from Quercus robur. USA, Aspergillus arizonicus from indoor air in a hospital, Caeliomyces tampanus (incl. Caeliomyces gen. nov.) from office dust, Cippumomyces mortalis (incl. Cippumomyces gen. nov.) from a tombstone, Cylindrium desperesense from air in a store, Tetracoccosporium pseudoaerium from air sample in house, Toxicocladosporium glendoranum from air in a brick room, Toxicocladosporium losalamitosense from air in a classroom, Valsonectria portsmouthensis from air in men’s locker room and Varicosporellopsis americana from sludge in a water reservoir. Vietnam, Entoloma kovalenkoi on rotten wood, Fusarium chuoi inside seed of Musa itinerans, Micropsalliota albofelina on soil in tropical evergreen mixed forests and Phytophthora docyniae from soil and roots of Docynia indica. Morphological and culture characteristics are supported by DNA barcodes.
... Lepiota sect. Lilaceae with a hymenoderm structure of the pileus covering is not monophyletic (Vellinga 2003(Vellinga , 2010. In phylogenetic analysis based on ITS and LSU (Fig. 23a, Etymology: the name 'pleurocystidiata' is from the presence of pleurocystidia. ...




The recent realistic estimate of fungal numbers which used various algorithms was between 2.2 and 3.8 million. There are nearly 100,000 accepted species of Fungi and fungus-like taxa, which is between 2.6 and 4.5% of the estimated species. Several forums such as Botanica Marina series, Fungal Diversity notes, Fungal Biodiversity Profiles, Fungal Systematics and Evolution—New and Interesting Fungi, Mycosphere notes and Fungal Planet have enhanced the introduction of new taxa and nearly 2000 species have been introduced in these publications in the last decade. The need to define a fungal species more accurately has been recognized, but there is much research needed before this can be better clarified. We address the evidence that is needed to estimate the numbers of fungi and address the various advances that have been made towards its understanding. Some genera are barely known, whereas some plant pathogens comprise numerous species complexes and numbers are steadily increasing. In this paper, we examine ten genera as case studies to establish trends in fungal description and introduce new species in each genus. The genera are the ascomycetes Colletotrichum and Pestalotiopsis (with many species or complexes), Atrocalyx, Dothiora, Lignosphaeria, Okeanomyces, Rhamphoriopsis, Thozetella, Thyrostroma (relatively poorly studied genera) and the basidiomycete genus Lepiota. We provide examples where knowledge is incomplete or lacking and suggest areas needing further research. These include (1) the need to establish what is a species, (2) the need to establish how host-specific fungi are, not in highly disturbed urban areas, but in pristine or relatively undisturbed forests, and (3) the need to establish if species in different continents, islands, countries or regions are different, or if the same fungi occur worldwide? Finally, we conclude whether we are anywhere near to flattening the curve in new species description.
... However, some species were placed in other sections of Lepiota because of their spurred spores (Bon 1981, Horak 1981, Candusso & Lanzoni 1990. Vellinga and Huijser (1998), and Vellinga (2001aVellinga ( , 2010 discussed the history of section Lilaceae and gave an overview of European and western North American species. Liang et al. (2009) investigated the species with spurred spores related to Lepiota cristata (Bolton) P. Kumm. ...





During our studies of the genus Lepiota in northern Thailand we collected a putatively new species with a distinct morphology and ITS nrDNA profile from Chiang Mai Province. The new species, Lepiota condylospora, is characterized by the presence of reddish brown to brownish orange or brown squamules on the pileus surface, triangular basidiospores with two lateral knobs, and a hymenidermal pileipellis composed of broadly to narrowly clavate elements. Two genetically distinct species of Lepiota section Lilaceae having triangular spores with two lateral knobs are compared with L. condylospora: Lepiota fraterna, from Papua New Guinea, differs in having larger basidiospores and cheilocystidia; while L. cristata var. macrospora, from China, has bigger basidiomata. A full description, color photographs, line drawings and a phylogenetic tree to show the position of the new species are provided.
... includes the pale-spored members of the Agaricaceae Chevall., which are circumscribed by having non-metachromatic, dextrinoid, and usually binucleate spores, cheilocystidia usually present, pleurocystidia absent, a regular hymenophoral trama, and clamp-connections usually present. The structure of the pileus covering has been shown to be a key character to divide the genus into operative, morphology-based sections (Vellinga and Huijser 1999;Vellinga 2001Vellinga , 2003Vellinga , 2010. ...
... Sequences obtained in this study were compared to those available in the GenBank (http://www.ncbi.nlm.nih.gov/) and UNITE (http://unite.ut.ee/) databases by using the Blastn algorithm (Altschul et al. 1990). Based on the BLASTn results (sequences were selected based on the greatest similarity) and outcomes of recent phylogenetic studies incorporating Lepiota sequences (Vellinga 2003(Vellinga , 2004(Vellinga , 2010Vizzini et al. 2014a, b;Justo et al. 2015;Qasim et al. 2015;Hosen et al. 2016) sequences were retrieved from GenBank for the comparative phylogenetic analysis. The nrITS and nrLSU datasets were analysed separately. ...
... The nrITS dataset was partitioned into ITS1, 5.8S and ITS2 subsets. Chamaemyces fracidus (AY176343 and AY176344) was used as an outgroup taxon in both the nrITS and nrLSU analyses because it is basal in the Agaricaceae (Vellinga 2004(Vellinga , 2010. ...



Lepiota psalion is fully described based on a recent collection from Sardinia (Italy) and the holotype. NrITS-and nrLSU-based phylogeny demonstrates that sequences deposited in GenBank as "L. psalion" and generated from two Dutch and one Chinese collections are not conspecific with the holotype and represent two distinct, undescribed species. These species are here proposed as Lepiota recondita sp. nov. and Lepiota sinorecondita ad int.
... PCR amplification, sequencing and sequence alignment described in the previous study of Hosen et al. (2013) were followed. Based on the BLASTn results against NCBI, and recent phylogenetic studies of lepiotaceous fungi (Vellinga 2003(Vellinga , 2010Liang et al. 2011;Nawaz et al. 2013;Vizzini et al. 2014;Justo et al. 2015;Qasim et al. 2016), a total of 57 ITS (ITS1-5.8S-ITS2) sequences were retrieved from GenBank, and then combined with the two newly generated ITS sequences of the proposed new species. ...
... Lepiota pyrochroa, a species reported from Europe (Netherlands and France), differs from the new species in having a slightly micaceous completely orange-red pileus surface where the elements are tightly knit together, and comparatively smaller basidiospores 4.4-5.5 × 3.0-3.7 µm (Malençon & Bertault 1970, Vellinga & Huijser 1999. Lepiota lilacea has a smaller pileus up to 36 mm broad, with a dark violet disc, a distinct annulus, and nondextrinoid basidiospores (Vellinga & Huijser 1999, Vellinga 2010). ...

Lepiota bengalensis is introduced as a new species based on material collected in Bangladesh. This species is characterized by its small to medium-sized basidiomata, brownish red, reddish brown to reddish orange squamules on pileus, fasciculate with narrowly clavate cheilocystidia, a hymeniform pileus covering, and strongly dextrinoid, cyanophilic, ellipsoid-cylindrical basidiospores. This species is a member of Lepiota sect. Lilaceae, and phylogenetically closely related to the European species L. ochraceofulva based on the nuclear ribosomal internal transcribed spacer (ITS) sequence data. Descriptions, color images of the basidiomata and line drawings are presented.
... these sequences mainly come from the studies of baroni et al. Sysouphanthong et al. (2011Sysouphanthong et al. ( , 2012, Vellinga (2001aVellinga ( , 2001bVellinga ( , 2001cVellinga ( , 2003Vellinga ( , 2004Vellinga ( , 2007Vellinga ( , 2010, Vellinga et al. (2011) and Vizzini et al. (2014aVizzini et al. ( , 2014b. Sequences were aligned using MAfft version 7 (Katoh & toh 2008) and the strategy fft-NS-i was selected. ...


Smithiomyces dominicanus is described as new, based on collections made in the Dominican Republic. Morphologically this taxon is very similar to S. mexicanus, but differs in the non-umbonate pileus, the broad stipe base and the smooth basidiospores as seen under light microscope, but with isolated, small warts under SEM. Molecular (nrITS) data also supports the recognition of this taxon as a separate species.