Figure 4 - uploaded by Pablo Billeke
Content may be subject to copyright.
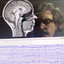
![Electrical brain activity in the expectancy phase. ( A ) Time – frequency charts of differences between HR and LR offers using dB unit (10 × [log 10 (HR) − log 10 (LR)]). The right panel shows the scalp distribution and the estimated source of power differences between conditions. ( B ) Trial-by-trial analysis using mixed linear model. Color represents the t -value of areas with signi fi cant correlation between the amplitude of early alpha band and logit of the probability of acceptances (logit( A )), and signi fi cant correlation between the amplitude of late alpha band and logit of the entropy (logit( E )). In all scalp distributions and source estimates, we show only signi fi cant clusters. See also Supplementary Figure 5 and Tables 6 – 9.](profile/Pablo-Billeke/publication/230766939/figure/fig4/AS:300560277360643@1448670562756/Electrical-brain-activity-in-the-expectancy-phase-A-Time-frequency-charts-of.png)
Electrical brain activity in the expectancy phase. ( A ) Time – frequency charts of differences between HR and LR offers using dB unit (10 × [log 10 (HR) − log 10 (LR)]). The right panel shows the scalp distribution and the estimated source of power differences between conditions. ( B ) Trial-by-trial analysis using mixed linear model. Color represents the t -value of areas with signi fi cant correlation between the amplitude of early alpha band and logit of the probability of acceptances (logit( A )), and signi fi cant correlation between the amplitude of late alpha band and logit of the entropy (logit( E )). In all scalp distributions and source estimates, we show only signi fi cant clusters. See also Supplementary Figure 5 and Tables 6 – 9.
Source publication









In social interactions, the perception of how risky our decisions are depends on how we anticipate other people's behaviors. We used electroencephalography to study the neurobiology of perception of social risk, in subjects playing the role of proposers in an iterated ultimatum game in pairs. Based on statistical modeling, we used the previous beha...
Contexts in source publication
Context 1
... and p3), we carried out 2 control analyses. In one we computed the MFN amplitude relative to neighboring positive components (( p3 + p2)/2 – MFN, UR = 0.253 μ V/cm 2 , ER = 0.173 μ V/cm 2 , P = 0.0012). In the other analysis, we fi xed the most negative point occurring in the time window 250 – 300 ms per subject to compute the mean across subjects ( P < 0.001). In both analyses, the MFN difference remained signi fi cant. In accordance with previous studies, the signi fi cant differences in the estimated cortical sources of this component were located in the mPFC and in the medial posterior region (Miltner et al. 1997; Gehring and Willoughby 2002; Luu et al. 2003; Polezzi et al. 2010). We then explored the time – frequency dynamics during the feedback phase and found that risk modulated activity in 2 frequency bands (Fig. 3). In frontal regions, between 240 and 400 ms after the feedback, URs presented a greater upper alpha band power (10 – 15 Hz) than that of ERs (main peak in FCz electrode, P = 0.0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). In a more extended time window (250 – 700 ms), theta band activity (4 – 7 Hz) was characterized by a stronger increase in power after URs (main peak in FCz electrode, P = 0.00004 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). We therefore focused on these time – frequency windows of interest for further analysis (see the Materials and Methods section). In agreement with previous reports (Christie and Tata 2009), the differences in the estimated cortical sources of theta activity were located in the mPFC, mainly on the right hemisphere, and in the medial posterior region. The sources of high alpha activity were located in the mPFC and lateral prefrontal cortex (lPFC; see also Supplementary Fig. 3 for comparison between different alpha ranges). Since expectations changed theta activity, we evaluated whether the degree of risk modulated activity in these frequency bands. For this end, we carried out a trial-by-trial analysis using a mixed linear regression. We estimated the degree of risk using both the probability of acceptance (using the logit transform of the probability of acceptance, logit( A )) and the Shannon entropy of the possible responses (logit( E )). In both cases, we used the coef fi cients of the logistic mixed model of the behavioral study (Table 1). Due to the fact that theta activity may correlate with both responses and expectations, we separated activity related to rejections from that related to acceptances and included the response as a factor in the model. In this analysis, logit( A ) signi fi cantly predicted theta power in rejections (main peaks in Fz and FCz electrodes, P < 0.01 corrected by FDR, Fig. 3 and see Supplementary Table 5). Interestingly, when we added the logit ( E ) (as a measure of the variability of the possible outcomes), it did not turn out to be a signi fi cant predictor. The source estimation of the logit( A ) coef fi cient in rejections was located in the right lPFC, mPFC, and medial posterior region and the source of the responses (as a factor) was located in mPFC and lPFC. In the case of acceptances, logit( A ) sources were placed in mPFC, but these did not survive the multiple comparison correction. All of these results were replicated using a wavelets transform, suggesting that our results are not an artifact of the analytical method (see Supplementary Fig. 4). None of these analyses was signi fi cant for upper alpha band activity. Since feedback-related brain activity changed under different risk perceptions, proposers should have had different anticipatory-related brain activity according to the way they estimated how risky the proposal they had just made was. In order to study this, we explored the period after proposers sent their offers but prior to receiving the corresponding answer from the responder. As in the feedback phase, we fi rst made an exploratory analysis of brain oscillatory activity. In right-central electrodes, we found clear differential anticipatory activity in alpha bands. Alpha activity presented a signi fi cant right-lateralized drop in HR offers in the 400 – 800 ms window after the offer was made (mean peaks in TP8 and FC4 electrodes, P < 0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 4 and see Supplementary Table 6). Brain sources estimated for this difference were distributed and included temporo-parietal regions, the left lPFC and the right insular cortex. We therefore focused on these time – frequency windows of interest in order to evaluate whether alpha power depended on risk. For this, we used logit( A ) and logit( E ) as measures of risk. Interestingly, trial-by-trial alpha power correlated positively with logit ( A ) (using a mixed linear model, main effect in FT8 and TP8 electrodes, P < 0.01 corrected by FDR, Fig. 4 and see Supplementary Table 7). Notably, unlike feedback theta activity, when we added the logit( E ) to the model, it also resulted in a signi fi cant predictor (see Supplementary Tables 8 and 9). Previous studies have shown speci fi c temporal dynamics of mesolimbic dopaminergic neurons related to the probabilistic reward anticipation. Some neurons present a transient activation that correlates with the probability of the reward, whereas other neurons present a tonic activation (increasing over the time interval), which is related to the variability or uncertainty of the reward (Schultz 1998; Schultz et al. 2008). To evaluate whether alpha activity has a similar temporal dynamics, we examined an additional late time window (late alpha, − 700 to − 200 ms before the feedback). Interestingly, early alpha fi tted best with the probability, while late alpha correlated better with the entropy (see Supplementary Table 9). In the brain source space, the correlation between early alpha and logit( A ) included the temporo-parietal regions and the insular cortex of the right hemisphere. Interestingly, the correlation between late alpha and logit( E ) included sources in the bilateral medial frontal cortex (Fig. 4). Like in the case of brain activity during the feedback phase, these results were also replicated using a wavelets transform (see Supplementary Fig. 5). As discussed previously, the proposer ’ s behavior was in fl u- enced by both the risk of the preceding offer and the corresponding responder ’ s feedback. This became evident in the change of the next offer in both experiments (Fig. 2 E and see Supplementary S1). Thus, each risk – response combination was followed by a particular change of offer (ER: $+8.1, UA: $ +4.3, UR: $ − 2.1 and EA: $ − 4.5; P < 0.001). Due to the fact that theta and alpha bands showed speci fi c risk modulations, they could underlie fi ne behavioral adaptation processes in social interactions. For example, prior reports suggest that frontal theta band signals error prediction in the context of behavioral adaptation (Cavanagh et al. 2010). To look into this possibility, we analyzed whether the power of these frequency bands predicted the next behavior in a trial-by-trial analysis. Importantly, feedback theta power and expectancy late alpha power together predicted signi fi cantly the next change of offer (alpha main peak TP8 electrode, P < 0.001 uncorrected; theta main effect in FCz and FP1 electrodes, P < 0.001 uncorrected; Fig. 5 and see Supplementary Tables 10 and 11). In accordance with an overall negotiating attitude, we found a strong negative correlation between the current change of offer and the next change of offer (Spearman ’ s ρ = − 0.41, P < 0.001). In other words, after an HR offer, the proposers were willing to make an LR offer and vice versa. This prompted us to include the current change of offer in the model. We were thus able to evaluate whether oscillatory activity underlies behavioral regulation necessary for this social negotiation and is not just an epiphe- nomenon of it. Interestingly, in this model, late alpha and feedback theta were signi fi cant predictors for the next change of offer. In the brain source space, late alpha prediction was associated with activity in the right temporo-parietal region and left insular cortex, and theta prediction was related to activity in lPFC, mPFC, and posterior cingulate cortex (PCC) for rejections. In the case of acceptances, the sources in mPFC did not survive the multiple comparison correction. As in previous analyses, results were replicated using wavelets transform (see Supplementary Fig. 6), con fi rming that our results where not biased by the use of the windowed fast-Fourier transform approach. Theta band activity, thereby, seems to re fl ect brain mechanisms that sense the prediction error: After an acceptance, proposers tend to decrease their offers and, indeed, in this case, theta activity correlated negatively with risk and predicted a smaller decrease in the next offer (Fig. 5; see Supplementary Table 11). In contrast, after a rejection, proposers tend to increase the offer. In this case, theta correlated positively with risk and predicted a smaller increase in the next offer. On the other hand, the fall in alpha band seems to re fl ect a prepara- tory activity for a possible rejection. Accordingly, it correlated with risk and predicted, only when a rejection occurred, an increase in the next offer. People participating in social interactions are able to distinguish safe from risky decisions. In making this distinction, people integrate others ’ intentions and the way others perceive their own intentions. In order to investigate how this integration occurs both at the behavioral and the neural levels, the current study examined individuals playing the iterated ultimatum game in a behavioral and an EEG experiment. The behavioral results indicate that responders reject offers that are far from the ...
Context 2
... taken in the next round. In the EEG study, subjects played as proposers against simulated partners whom they believed were human. We used the coef fi cients of the logistic mixed model to simulate responders (see the Materials and Methods section). The distributions of acceptances and rejections, and the offering behaviors related to a rejection were similar to the corresponding ones in the behavioral study (Fig. 2 and see Supplementary Fig. 1), suggesting that simulated responders elicited comparable behaviors in proposers. If our risk classi fi cation was consistent with the risk perceived by the proposer, the error-related activity evoked by the unexpected responses should be larger than that evoked by the expected responses. Speci fi cally, when proposers perceive LRs for their offers, rejections should elicit a higher MFN than when they perceive HRs. To test this hypothesis, we analyzed the time window when a proposer received a rejection. As conjectured, we found a signi fi cant difference between the MFN component of URs and that of ERs in fronto-medial electrodes (main peak in FCz electrode, P = 0.00003 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, see Fig. 3 and Supplementary Table 3 and Fig. 3 for the time course of the evoked response over all scalp electrodes). To rule out the possibility that this difference was due to differences arising from neighboring positive components (mainly p2 and p3), we carried out 2 control analyses. In one we computed the MFN amplitude relative to neighboring positive components (( p3 + p2)/2 – MFN, UR = 0.253 μ V/cm 2 , ER = 0.173 μ V/cm 2 , P = 0.0012). In the other analysis, we fi xed the most negative point occurring in the time window 250 – 300 ms per subject to compute the mean across subjects ( P < 0.001). In both analyses, the MFN difference remained signi fi cant. In accordance with previous studies, the signi fi cant differences in the estimated cortical sources of this component were located in the mPFC and in the medial posterior region (Miltner et al. 1997; Gehring and Willoughby 2002; Luu et al. 2003; Polezzi et al. 2010). We then explored the time – frequency dynamics during the feedback phase and found that risk modulated activity in 2 frequency bands (Fig. 3). In frontal regions, between 240 and 400 ms after the feedback, URs presented a greater upper alpha band power (10 – 15 Hz) than that of ERs (main peak in FCz electrode, P = 0.0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). In a more extended time window (250 – 700 ms), theta band activity (4 – 7 Hz) was characterized by a stronger increase in power after URs (main peak in FCz electrode, P = 0.00004 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). We therefore focused on these time – frequency windows of interest for further analysis (see the Materials and Methods section). In agreement with previous reports (Christie and Tata 2009), the differences in the estimated cortical sources of theta activity were located in the mPFC, mainly on the right hemisphere, and in the medial posterior region. The sources of high alpha activity were located in the mPFC and lateral prefrontal cortex (lPFC; see also Supplementary Fig. 3 for comparison between different alpha ranges). Since expectations changed theta activity, we evaluated whether the degree of risk modulated activity in these frequency bands. For this end, we carried out a trial-by-trial analysis using a mixed linear regression. We estimated the degree of risk using both the probability of acceptance (using the logit transform of the probability of acceptance, logit( A )) and the Shannon entropy of the possible responses (logit( E )). In both cases, we used the coef fi cients of the logistic mixed model of the behavioral study (Table 1). Due to the fact that theta activity may correlate with both responses and expectations, we separated activity related to rejections from that related to acceptances and included the response as a factor in the model. In this analysis, logit( A ) signi fi cantly predicted theta power in rejections (main peaks in Fz and FCz electrodes, P < 0.01 corrected by FDR, Fig. 3 and see Supplementary Table 5). Interestingly, when we added the logit ( E ) (as a measure of the variability of the possible outcomes), it did not turn out to be a signi fi cant predictor. The source estimation of the logit( A ) coef fi cient in rejections was located in the right lPFC, mPFC, and medial posterior region and the source of the responses (as a factor) was located in mPFC and lPFC. In the case of acceptances, logit( A ) sources were placed in mPFC, but these did not survive the multiple comparison correction. All of these results were replicated using a wavelets transform, suggesting that our results are not an artifact of the analytical method (see Supplementary Fig. 4). None of these analyses was signi fi cant for upper alpha band activity. Since feedback-related brain activity changed under different risk perceptions, proposers should have had different anticipatory-related brain activity according to the way they estimated how risky the proposal they had just made was. In order to study this, we explored the period after proposers sent their offers but prior to receiving the corresponding answer from the responder. As in the feedback phase, we fi rst made an exploratory analysis of brain oscillatory activity. In right-central electrodes, we found clear differential anticipatory activity in alpha bands. Alpha activity presented a signi fi cant right-lateralized drop in HR offers in the 400 – 800 ms window after the offer was made (mean peaks in TP8 and FC4 electrodes, P < 0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 4 and see Supplementary Table 6). Brain sources estimated for this difference were distributed and included temporo-parietal regions, the left lPFC and the right insular cortex. We therefore focused on these time – frequency windows of interest in order to evaluate whether alpha power depended on risk. For this, we used logit( A ) and logit( E ) as measures of risk. Interestingly, trial-by-trial alpha power correlated positively with logit ( A ) (using a mixed linear model, main effect in FT8 and TP8 electrodes, P < 0.01 corrected by FDR, Fig. 4 and see Supplementary Table 7). Notably, unlike feedback theta activity, when we added the logit( E ) to the model, it also resulted in a signi fi cant predictor (see Supplementary Tables 8 and 9). Previous studies have shown speci fi c temporal dynamics of mesolimbic dopaminergic neurons related to the probabilistic reward anticipation. Some neurons present a transient activation that correlates with the probability of the reward, whereas other neurons present a tonic activation (increasing over the time interval), which is related to the variability or uncertainty of the reward (Schultz 1998; Schultz et al. 2008). To evaluate whether alpha activity has a similar temporal dynamics, we examined an additional late time window (late alpha, − 700 to − 200 ms before the feedback). Interestingly, early alpha fi tted best with the probability, while late alpha correlated better with the entropy (see Supplementary Table 9). In the brain source space, the correlation between early alpha and logit( A ) included the temporo-parietal regions and the insular cortex of the right hemisphere. Interestingly, the correlation between late alpha and logit( E ) included sources in the bilateral medial frontal cortex (Fig. 4). Like in the case of brain activity during the feedback phase, these results were also replicated using a wavelets transform (see Supplementary Fig. 5). As discussed previously, the proposer ’ s behavior was in fl u- enced by both the risk of the preceding offer and the corresponding responder ’ s feedback. This became evident in the change of the next offer in both experiments (Fig. 2 E and see Supplementary S1). Thus, each risk – response combination was followed by a particular change of offer (ER: $+8.1, UA: $ +4.3, UR: $ − 2.1 and EA: $ − 4.5; P < 0.001). Due to the fact that theta and alpha bands showed speci fi c risk modulations, they could underlie fi ne behavioral adaptation processes in social interactions. For example, prior reports suggest that frontal theta band signals error prediction in the context of behavioral adaptation (Cavanagh et al. 2010). To look into this possibility, we analyzed whether the power of these frequency bands predicted the next behavior in a trial-by-trial analysis. Importantly, feedback theta power and expectancy late alpha power together predicted signi fi cantly the next change of offer (alpha main peak TP8 electrode, P < 0.001 uncorrected; theta main effect in FCz and FP1 electrodes, P < 0.001 uncorrected; Fig. 5 and see Supplementary Tables 10 and 11). In accordance with an overall negotiating attitude, we found a strong negative correlation between the current change of offer and the next change of offer (Spearman ’ s ρ = − 0.41, P < 0.001). In other words, after an HR offer, the proposers were willing to make an LR offer and vice versa. This prompted us to include the current change of offer in the model. We were thus able to evaluate whether oscillatory activity underlies behavioral regulation necessary for this social negotiation and is not just an epiphe- nomenon of it. Interestingly, in this model, late alpha and feedback theta were signi fi cant predictors for the next change of offer. In the brain source space, late alpha prediction was associated with activity in the right temporo-parietal region and left insular cortex, and theta prediction was related to activity in lPFC, mPFC, and posterior cingulate cortex (PCC) for rejections. In the case of acceptances, the sources in mPFC did not survive the multiple ...
Context 3
... the time window when a proposer received a rejection. As conjectured, we found a signi fi cant difference between the MFN component of URs and that of ERs in fronto-medial electrodes (main peak in FCz electrode, P = 0.00003 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, see Fig. 3 and Supplementary Table 3 and Fig. 3 for the time course of the evoked response over all scalp electrodes). To rule out the possibility that this difference was due to differences arising from neighboring positive components (mainly p2 and p3), we carried out 2 control analyses. In one we computed the MFN amplitude relative to neighboring positive components (( p3 + p2)/2 – MFN, UR = 0.253 μ V/cm 2 , ER = 0.173 μ V/cm 2 , P = 0.0012). In the other analysis, we fi xed the most negative point occurring in the time window 250 – 300 ms per subject to compute the mean across subjects ( P < 0.001). In both analyses, the MFN difference remained signi fi cant. In accordance with previous studies, the signi fi cant differences in the estimated cortical sources of this component were located in the mPFC and in the medial posterior region (Miltner et al. 1997; Gehring and Willoughby 2002; Luu et al. 2003; Polezzi et al. 2010). We then explored the time – frequency dynamics during the feedback phase and found that risk modulated activity in 2 frequency bands (Fig. 3). In frontal regions, between 240 and 400 ms after the feedback, URs presented a greater upper alpha band power (10 – 15 Hz) than that of ERs (main peak in FCz electrode, P = 0.0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). In a more extended time window (250 – 700 ms), theta band activity (4 – 7 Hz) was characterized by a stronger increase in power after URs (main peak in FCz electrode, P = 0.00004 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). We therefore focused on these time – frequency windows of interest for further analysis (see the Materials and Methods section). In agreement with previous reports (Christie and Tata 2009), the differences in the estimated cortical sources of theta activity were located in the mPFC, mainly on the right hemisphere, and in the medial posterior region. The sources of high alpha activity were located in the mPFC and lateral prefrontal cortex (lPFC; see also Supplementary Fig. 3 for comparison between different alpha ranges). Since expectations changed theta activity, we evaluated whether the degree of risk modulated activity in these frequency bands. For this end, we carried out a trial-by-trial analysis using a mixed linear regression. We estimated the degree of risk using both the probability of acceptance (using the logit transform of the probability of acceptance, logit( A )) and the Shannon entropy of the possible responses (logit( E )). In both cases, we used the coef fi cients of the logistic mixed model of the behavioral study (Table 1). Due to the fact that theta activity may correlate with both responses and expectations, we separated activity related to rejections from that related to acceptances and included the response as a factor in the model. In this analysis, logit( A ) signi fi cantly predicted theta power in rejections (main peaks in Fz and FCz electrodes, P < 0.01 corrected by FDR, Fig. 3 and see Supplementary Table 5). Interestingly, when we added the logit ( E ) (as a measure of the variability of the possible outcomes), it did not turn out to be a signi fi cant predictor. The source estimation of the logit( A ) coef fi cient in rejections was located in the right lPFC, mPFC, and medial posterior region and the source of the responses (as a factor) was located in mPFC and lPFC. In the case of acceptances, logit( A ) sources were placed in mPFC, but these did not survive the multiple comparison correction. All of these results were replicated using a wavelets transform, suggesting that our results are not an artifact of the analytical method (see Supplementary Fig. 4). None of these analyses was signi fi cant for upper alpha band activity. Since feedback-related brain activity changed under different risk perceptions, proposers should have had different anticipatory-related brain activity according to the way they estimated how risky the proposal they had just made was. In order to study this, we explored the period after proposers sent their offers but prior to receiving the corresponding answer from the responder. As in the feedback phase, we fi rst made an exploratory analysis of brain oscillatory activity. In right-central electrodes, we found clear differential anticipatory activity in alpha bands. Alpha activity presented a signi fi cant right-lateralized drop in HR offers in the 400 – 800 ms window after the offer was made (mean peaks in TP8 and FC4 electrodes, P < 0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 4 and see Supplementary Table 6). Brain sources estimated for this difference were distributed and included temporo-parietal regions, the left lPFC and the right insular cortex. We therefore focused on these time – frequency windows of interest in order to evaluate whether alpha power depended on risk. For this, we used logit( A ) and logit( E ) as measures of risk. Interestingly, trial-by-trial alpha power correlated positively with logit ( A ) (using a mixed linear model, main effect in FT8 and TP8 electrodes, P < 0.01 corrected by FDR, Fig. 4 and see Supplementary Table 7). Notably, unlike feedback theta activity, when we added the logit( E ) to the model, it also resulted in a signi fi cant predictor (see Supplementary Tables 8 and 9). Previous studies have shown speci fi c temporal dynamics of mesolimbic dopaminergic neurons related to the probabilistic reward anticipation. Some neurons present a transient activation that correlates with the probability of the reward, whereas other neurons present a tonic activation (increasing over the time interval), which is related to the variability or uncertainty of the reward (Schultz 1998; Schultz et al. 2008). To evaluate whether alpha activity has a similar temporal dynamics, we examined an additional late time window (late alpha, − 700 to − 200 ms before the feedback). Interestingly, early alpha fi tted best with the probability, while late alpha correlated better with the entropy (see Supplementary Table 9). In the brain source space, the correlation between early alpha and logit( A ) included the temporo-parietal regions and the insular cortex of the right hemisphere. Interestingly, the correlation between late alpha and logit( E ) included sources in the bilateral medial frontal cortex (Fig. 4). Like in the case of brain activity during the feedback phase, these results were also replicated using a wavelets transform (see Supplementary Fig. 5). As discussed previously, the proposer ’ s behavior was in fl u- enced by both the risk of the preceding offer and the corresponding responder ’ s feedback. This became evident in the change of the next offer in both experiments (Fig. 2 E and see Supplementary S1). Thus, each risk – response combination was followed by a particular change of offer (ER: $+8.1, UA: $ +4.3, UR: $ − 2.1 and EA: $ − 4.5; P < 0.001). Due to the fact that theta and alpha bands showed speci fi c risk modulations, they could underlie fi ne behavioral adaptation processes in social interactions. For example, prior reports suggest that frontal theta band signals error prediction in the context of behavioral adaptation (Cavanagh et al. 2010). To look into this possibility, we analyzed whether the power of these frequency bands predicted the next behavior in a trial-by-trial analysis. Importantly, feedback theta power and expectancy late alpha power together predicted signi fi cantly the next change of offer (alpha main peak TP8 electrode, P < 0.001 uncorrected; theta main effect in FCz and FP1 electrodes, P < 0.001 uncorrected; Fig. 5 and see Supplementary Tables 10 and 11). In accordance with an overall negotiating attitude, we found a strong negative correlation between the current change of offer and the next change of offer (Spearman ’ s ρ = − 0.41, P < 0.001). In other words, after an HR offer, the proposers were willing to make an LR offer and vice versa. This prompted us to include the current change of offer in the model. We were thus able to evaluate whether oscillatory activity underlies behavioral regulation necessary for this social negotiation and is not just an epiphe- nomenon of it. Interestingly, in this model, late alpha and feedback theta were signi fi cant predictors for the next change of offer. In the brain source space, late alpha prediction was associated with activity in the right temporo-parietal region and left insular cortex, and theta prediction was related to activity in lPFC, mPFC, and posterior cingulate cortex (PCC) for rejections. In the case of acceptances, the sources in mPFC did not survive the multiple comparison correction. As in previous analyses, results were replicated using wavelets transform (see Supplementary Fig. 6), con fi rming that our results where not biased by the use of the windowed fast-Fourier transform approach. Theta band activity, thereby, seems to re fl ect brain mechanisms that sense the prediction error: After an acceptance, proposers tend to decrease their offers and, indeed, in this case, theta activity correlated negatively with risk and predicted a smaller decrease in the next offer (Fig. 5; see Supplementary Table 11). In contrast, after a rejection, proposers tend to increase the offer. In this case, theta correlated positively with risk and predicted a smaller increase in the next offer. On the other hand, the fall in alpha band seems to re fl ect a prepara- tory activity for a possible rejection. Accordingly, it correlated with risk and predicted, only when a ...
Context 4
... by a stronger increase in power after URs (main peak in FCz electrode, P = 0.00004 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 3 and see Supplementary Table 4). We therefore focused on these time – frequency windows of interest for further analysis (see the Materials and Methods section). In agreement with previous reports (Christie and Tata 2009), the differences in the estimated cortical sources of theta activity were located in the mPFC, mainly on the right hemisphere, and in the medial posterior region. The sources of high alpha activity were located in the mPFC and lateral prefrontal cortex (lPFC; see also Supplementary Fig. 3 for comparison between different alpha ranges). Since expectations changed theta activity, we evaluated whether the degree of risk modulated activity in these frequency bands. For this end, we carried out a trial-by-trial analysis using a mixed linear regression. We estimated the degree of risk using both the probability of acceptance (using the logit transform of the probability of acceptance, logit( A )) and the Shannon entropy of the possible responses (logit( E )). In both cases, we used the coef fi cients of the logistic mixed model of the behavioral study (Table 1). Due to the fact that theta activity may correlate with both responses and expectations, we separated activity related to rejections from that related to acceptances and included the response as a factor in the model. In this analysis, logit( A ) signi fi cantly predicted theta power in rejections (main peaks in Fz and FCz electrodes, P < 0.01 corrected by FDR, Fig. 3 and see Supplementary Table 5). Interestingly, when we added the logit ( E ) (as a measure of the variability of the possible outcomes), it did not turn out to be a signi fi cant predictor. The source estimation of the logit( A ) coef fi cient in rejections was located in the right lPFC, mPFC, and medial posterior region and the source of the responses (as a factor) was located in mPFC and lPFC. In the case of acceptances, logit( A ) sources were placed in mPFC, but these did not survive the multiple comparison correction. All of these results were replicated using a wavelets transform, suggesting that our results are not an artifact of the analytical method (see Supplementary Fig. 4). None of these analyses was signi fi cant for upper alpha band activity. Since feedback-related brain activity changed under different risk perceptions, proposers should have had different anticipatory-related brain activity according to the way they estimated how risky the proposal they had just made was. In order to study this, we explored the period after proposers sent their offers but prior to receiving the corresponding answer from the responder. As in the feedback phase, we fi rst made an exploratory analysis of brain oscillatory activity. In right-central electrodes, we found clear differential anticipatory activity in alpha bands. Alpha activity presented a signi fi cant right-lateralized drop in HR offers in the 400 – 800 ms window after the offer was made (mean peaks in TP8 and FC4 electrodes, P < 0001 uncorrected, and P < 0.001 corrected by the cluster-based permutation test, Fig. 4 and see Supplementary Table 6). Brain sources estimated for this difference were distributed and included temporo-parietal regions, the left lPFC and the right insular cortex. We therefore focused on these time – frequency windows of interest in order to evaluate whether alpha power depended on risk. For this, we used logit( A ) and logit( E ) as measures of risk. Interestingly, trial-by-trial alpha power correlated positively with logit ( A ) (using a mixed linear model, main effect in FT8 and TP8 electrodes, P < 0.01 corrected by FDR, Fig. 4 and see Supplementary Table 7). Notably, unlike feedback theta activity, when we added the logit( E ) to the model, it also resulted in a signi fi cant predictor (see Supplementary Tables 8 and 9). Previous studies have shown speci fi c temporal dynamics of mesolimbic dopaminergic neurons related to the probabilistic reward anticipation. Some neurons present a transient activation that correlates with the probability of the reward, whereas other neurons present a tonic activation (increasing over the time interval), which is related to the variability or uncertainty of the reward (Schultz 1998; Schultz et al. 2008). To evaluate whether alpha activity has a similar temporal dynamics, we examined an additional late time window (late alpha, − 700 to − 200 ms before the feedback). Interestingly, early alpha fi tted best with the probability, while late alpha correlated better with the entropy (see Supplementary Table 9). In the brain source space, the correlation between early alpha and logit( A ) included the temporo-parietal regions and the insular cortex of the right hemisphere. Interestingly, the correlation between late alpha and logit( E ) included sources in the bilateral medial frontal cortex (Fig. 4). Like in the case of brain activity during the feedback phase, these results were also replicated using a wavelets transform (see Supplementary Fig. 5). As discussed previously, the proposer ’ s behavior was in fl u- enced by both the risk of the preceding offer and the corresponding responder ’ s feedback. This became evident in the change of the next offer in both experiments (Fig. 2 E and see Supplementary S1). Thus, each risk – response combination was followed by a particular change of offer (ER: $+8.1, UA: $ +4.3, UR: $ − 2.1 and EA: $ − 4.5; P < 0.001). Due to the fact that theta and alpha bands showed speci fi c risk modulations, they could underlie fi ne behavioral adaptation processes in social interactions. For example, prior reports suggest that frontal theta band signals error prediction in the context of behavioral adaptation (Cavanagh et al. 2010). To look into this possibility, we analyzed whether the power of these frequency bands predicted the next behavior in a trial-by-trial analysis. Importantly, feedback theta power and expectancy late alpha power together predicted signi fi cantly the next change of offer (alpha main peak TP8 electrode, P < 0.001 uncorrected; theta main effect in FCz and FP1 electrodes, P < 0.001 uncorrected; Fig. 5 and see Supplementary Tables 10 and 11). In accordance with an overall negotiating attitude, we found a strong negative correlation between the current change of offer and the next change of offer (Spearman ’ s ρ = − 0.41, P < 0.001). In other words, after an HR offer, the proposers were willing to make an LR offer and vice versa. This prompted us to include the current change of offer in the model. We were thus able to evaluate whether oscillatory activity underlies behavioral regulation necessary for this social negotiation and is not just an epiphe- nomenon of it. Interestingly, in this model, late alpha and feedback theta were signi fi cant predictors for the next change of offer. In the brain source space, late alpha prediction was associated with activity in the right temporo-parietal region and left insular cortex, and theta prediction was related to activity in lPFC, mPFC, and posterior cingulate cortex (PCC) for rejections. In the case of acceptances, the sources in mPFC did not survive the multiple comparison correction. As in previous analyses, results were replicated using wavelets transform (see Supplementary Fig. 6), con fi rming that our results where not biased by the use of the windowed fast-Fourier transform approach. Theta band activity, thereby, seems to re fl ect brain mechanisms that sense the prediction error: After an acceptance, proposers tend to decrease their offers and, indeed, in this case, theta activity correlated negatively with risk and predicted a smaller decrease in the next offer (Fig. 5; see Supplementary Table 11). In contrast, after a rejection, proposers tend to increase the offer. In this case, theta correlated positively with risk and predicted a smaller increase in the next offer. On the other hand, the fall in alpha band seems to re fl ect a prepara- tory activity for a possible rejection. Accordingly, it correlated with risk and predicted, only when a rejection occurred, an increase in the next offer. People participating in social interactions are able to distinguish safe from risky decisions. In making this distinction, people integrate others ’ intentions and the way others perceive their own intentions. In order to investigate how this integration occurs both at the behavioral and the neural levels, the current study examined individuals playing the iterated ultimatum game in a behavioral and an EEG experiment. The behavioral results indicate that responders reject offers that are far from the social norm of 50%, although there are important interindividual variations. In our iterated version of the ultimatum game, prior behaviors of both players are the only way to infer both the other ’ s intentions and how the other perceives his/her partner ’ s intentions. Notably, previous behavior in fl uences signi fi cantly the responders ’ responses, showing less interindividual variations than the in fl uence of the absolute offer made. We conjecture that within the social norm compliance generated whenever punishment is possible (Fehr and Fischbancher 2004; Spitzer et al. 2007), prior behavior exerts a fi ne-tuning regulation on both risk perception and the expectation of subsequent behavior in an iterated interaction. Accordingly, we have proposed a classi fi cation based on preceding behavior in order to separate HR offers from LR offers. This classi fi cation not only distinguishes offers with high rejection probability and high variability (both cri- teria that de fi ne risk), but also particular situations in a social interaction that generate distinct subsequent behaviors. We used the MFN, an ERP component that has been related to error predictions (Holroyd and Coles 2002), as a neuronal marker ...
Citations
... 14,28 Neurophysiological studies have remarked on the role of oscillatory activity in theta range in several aspects of cognitive control, including its deficit in pathologies. 7,[29][30][31][32][33][34] Oscillatory activity in the theta band over the frontal midline is associated with detecting, communicating, and implementing cognitive control. 29,35 Theta-band oscillations have been tied to adaptive control mechanisms during response conflict. ...


Adapting our behavior to environmental demands relies on our capacity to perceive and manage potential conflicts within our surroundings. While evidence implicates the involvement of the lateral prefrontal cortex and theta oscillations in detecting conflict stimuli, their roles in conflict expectation remain elusive. Consequently, the exact computations and neural mechanisms underlying these cognitive processes still need to be determined. To address this gap, we employed an integrative approach involving cognitive computational modeling, fMRI, TMS, and EEG. Our results revealed a computational process underlying conflict expectation, which correlated with activity in the superior frontal gyrus (SFG). Furthermore, rhythmic TMS in the theta range applied over the SFG, but not over the inferior frontal junction, induced endogenous theta activity, enhancing computations associated with conflict expectation. These findings provide compelling evidence for the causal involvement of SFG theta activity in learning and allocating cognitive resources to address forthcoming conflict stimuli.
Significant Statement
Alterations in the processing of expectations of conflict events have been associated with several neuropsychiatric disorders that significantly affect the quality of life for many individuals. This article describes a cognitive computation underlying the conflict expectation and its causal neural mechanism involving theta brain activity in the superior frontal gyrus (SFG). Thus, unraveling this mechanism holds promise for developing interventions to address cognitive alterations related to anticipation of conflict events in neuropsychiatric disorders, improving overall cognitive function and quality of life.
... After a decision, research has revealed sustained connectivity between the parietal region and prefrontal cortex, which influences future decision-making [19,20]. Unexpected outcomes in uncertain situations have been linked to prefrontal activity, as indicated by fMRI studies [11] and oscillatory activity from EEG recordings [21,22]. A large body of work has shown a correlation between frontal delta and theta activity and prediction errors in uncertain situations [21,[23][24][25]. ...
... We anticipated that the prediction error signal of the probability calculated with τ i = 0, representing a lower degree of uncertainty in decision-making, would be particularly affected by parietal TMS stimulation. For this, we explored frontal electrodes where oscillatory activity related to prediction error has been described in prior work [21][22][23]33]. ...
... Prefrontal oscillatory activity has a widely studied role in cognitive control and working memory [25,[53][54][55]. Extensive research has shown that prefrontal delta and theta activities correlate with prediction errors [18,[21][22][23][56][57][58][59]. Prior evidence showed that parietal and frontal areas sustain effective connectivity during and after a decision, and such activity biases follow decision-making [19,20]. ...
Humans often face the challenge of making decisions between ambiguous options. The level of ambiguity in decision-making has been linked to activity in the parietal cortex, but its exact computational role remains elusive. To test the hypothesis that the parietal cortex plays a causal role in computing ambiguous probabilities, we conducted consecutive fMRI and TMS-EEG studies. We found that participants assigned unknown probabilities to objective probabilities, elevating the uncertainty of their decisions. Parietal cortex activity correlated with the objective degree of ambiguity and with a process that underestimates the uncertainty during decision-making. Conversely, the midcingulate cortex (MCC) encodes prediction errors and increases its connectivity with the parietal cortex during outcome processing. Disruption of the parietal activity increased the uncertainty evaluation of the options, decreasing cingulate cortex oscillations during outcome evaluation and lateral frontal oscillations related to value ambiguous probability. These results provide evidence for a causal role of the parietal cortex in computing uncertainty during ambiguous decisions made by humans.
... For example, a hyperscanning study found that negative external feedback may induce a decline in the inter-brain synchrony in the low-frequency bands, suggesting that low frequency-bands like theta may be an important marker of social cognition and emotional engagement (Balconi et al., 2018a). Indeed, results from other studies also suggested that the frontal theta activity may be associated with strategic regulation of social information processing and emotional features in a social context (Billeke et al., 2013(Billeke et al., , 2014Cristofori et al., 2013). Therefore, in the current study, we focused on the role of mindfulness practices in the inter-brain synchrony of theta-band between participants in the feedback processing during a cooperative task. ...
... The study found that greater theta inter-brain synchrony was observed in the mindfulness group than in the non-mindfulness group in the processing of the successful and the failure feedbacks during cooperation. Previous research suggested that the theta band is associated with social empathy and cognitive control (Balconi et al., 2015;Billeke et al., 2013). The increased inter-brain synchrony may reflect a better social understanding and empathy among interacting individuals (Cui et al., 2012). ...


Mindfulness appears to improve empathy and understanding in relationships, which are necessary for successful cooperation. However, the impact of mindfulness on cooperation has not been fully studied. This study used hyperscanning technique to examine the effect of mindfulness on the inter-brain synchrony of interacting individuals during the cooperative tasks. Forty-one dyads were randomly assigned to a mindfulness group or a non-mindfulness group. Dyads of the mindfulness group performed a short mindfulness exercise following a 15-minute mindfulness audio guidance. Dyads of the non-mindfulness group were instructed to rest quietly with their eyes closed. Then, simultaneously and continuously EEG was recorded from all dyads when they completed a computer-based cooperative game task. Reaction times (RTs) and success rates were used to indicate the behavioral performance, and phase locking value (PLV) was used to indicate the inter-brain synchrony. The results showed that (1) Greater theta inter-brain synchrony during the cooperative computer game tasks was observed in the mindfulness group than in the non-mindfulness group; (2) Greater theta inter-brain synchrony was observed in the successful cooperation conditions as compared to those in the failure cooperation conditions; (3) Greater theta inter-brain synchrony was observed at the frontal region as compared to those at the parietal-occipital region in the successful cooperation condition. The results expand the neural basis of the effects of mindfulness on cooperation feedback processing.
... For instance, increased frontal theta activity has been related to observing others in pain (compared to neutral stimuli) as well as to one's own unpleasant feelings (26,27). Frontal theta activity is also related to self-reflexive thinking within social interactions (28) and predicting others' decisions (20,29). These data support the relevance of the presence of others in people's decision-making and that there are different ways in which others are included in the variables that a decision-maker takes into account while deciding. ...


Introduction
Decision-making is a process that can be strongly affected by social factors. Evidence has shown how people deviate from traditional rational-choice predictions under different levels of social interactions. The emergence of prosocial decision-making, defined as any action that is addressed to benefit another individual even at the expense of personal benefits, has been reported as an example of such social influence. Furthermore, brain evidence has shown the involvement of structures such as the prefrontal cortex, anterior insula, and midcingulate cortex during decision settings in which a decision maker interacts with others under physical pain or distress or while being observed by others.
Methods
Using a slightly modified version of the dictator game and EEG recordings, we tested the hypothesis that the inclusion of another person into the decision setting increases prosocial decisions in young adults and that this increase is higher when the other person is associated with others in need. At the brain level, we hypothesized that the increase in prosocial decisions correlates with frontal theta activity.
Results and Discussion
The results showed that including another person in the decision, setting increased prosocial behavior only when this presence was associated with someone in need. This effect was associated with an increase in frontocentral theta-oscillatory activity. These results suggest that the presence of someone in need enhances empathy concerns and norm compliance, raising the participants’ prosocial decision-making.
... After a decision, research has revealed sustained connectivity between the parietal region and prefrontal cortex, which influences future decision-making [19,20]. Unexpected outcomes in uncertain situations have been linked to prefrontal activity, as indicated by fMRI studies [11] and oscillatory activity from EEG recordings [21,22]. A large body of work has shown a correlation between frontal delta and theta activity and prediction errors in uncertain situations [21,[23][24][25]. ...
... We anticipated that the prediction error signal of the probability calculated with τi = 0, representing a lower degree of uncertainty in decisionmaking, would be particularly affected by parietal TMS stimulation. For this, we explored frontal electrodes where oscillatory activity related to prediction error has been described in prior work [21][22][23]33]. ...
... Prefrontal oscillatory activity has a widely studied role in cognitive control and working memory [25,[53][54][55]. Extensive research has shown that prefrontal delta and theta activities correlate with prediction errors [18,[21][22][23][56][57][58][59]. Prior evidence showed that parietal and frontal areas sustain effective connectivity during and after a decision, and such activity biases follow decision-making [19,20]. ...
Humans often face the challenge of making decisions between ambiguous options. The level of
ambiguity in decision-making has been linked to activity in the parietal cortex, but its exact
computational role remains elusive. To test the hypothesis that the parietal cortex plays a causal role
in computing ambiguous probabilities, we conducted consecutive fMRI and TMS-EEG studies. We
found that participants assigned unknown probabilities to objective probabilities, elevating the
uncertainty of their decisions. Parietal cortex activity correlated with the objective degree of
ambiguity and with a process that underestimates the uncertainty during decision-making.
Conversely, the midcingulate cortex encodes prediction errors and increases its connectivity with the
parietal cortex during outcome processing. Disruption of the parietal activity increased the uncertainty
evaluation of the options, decreasing cingulate cortex oscillations during outcome evaluation and
lateral frontal oscillations related to value ambiguous probability. These results provide evidence for
a causal role of the parietal cortex in computing uncertainty during ambiguous decisions made by
humans.
... After a decision, research has revealed sustained connectivity between the parietal region and prefrontal cortex, which influences future decision-making (19,20). Unexpected outcomes in uncertain situations have been linked to prefrontal activity, as indicated by fMRI studies (11) and oscillatory activity from EEG recordings (21,22). A large body of work has shown a correlation between frontal delta and theta activity and prediction errors in uncertain situations (21,(23)(24)(25). ...
... We anticipated that the prediction error signal of the probability calculated with , representing a lower degree of uncertainty in decision-making, would be particularly affected by parietal TMS stimulation. For this, we explored frontal electrodes where oscillatory activity related to prediction error has been described in prior work (21)(22)(23)31). ...
... Prefrontal oscillatory activity has a widely studied role in cognitive control and working memory (25,(50)(51)(52). Extensive research has shown that prefrontal delta and theta activity correlates with prediction errors (18,(21)(22)(23)(53)(54)(55)(56). Prior evidence showed that parietal and frontal areas sustain effective connectivity during and after a decision, and such activity biases follow decision-making (19,20). ...
Humans often face the challenge of making decisions between ambiguous options. The level of ambiguity in decision-making has been linked to activity in the parietal cortex, but its exact computational role remains elusive. To test the hypothesis that the parietal cortex plays a causal role in computing ambiguous probabilities, we conducted consecutive fMRI and TMS-EEG studies. We found that participants assigned unknown probabilities to objective probabilities, elevating the uncertainty of their decisions. Parietal cortex activity correlated with both the objective degree of ambiguity and a process that underestimates the uncertainty during decision-making. Conversely, the midcingulate cortex encodes prediction errors and increases its connectivity with the parietal cortex during outcome processing. Disruption of the parietal activity increased the uncertainty evaluation of the options, decreasing cingulate cortex oscillations during outcome evaluation. These results provide evidence for a causal role of the parietal cortex in computing uncertainty during ambiguous decisions made by humans.
... For instance, increased frontal theta activity has been related to observing others in pain (compared to neutral stimuli) as well as to one's own unpleasant feelings 21,22 . Frontal theta activity has also been related to self-re exive thinking within social interactions 23 and the prediction of others' decisions 16,24 . Taken together, these data support the relevance of the presence of others on people's decision making and that there are different ways in which a given other sneaks into the variables that a decision-maker takes into account while deciding. ...
Decision making is a process that can be strongly affected by social factors. Profuse evidence has shown how people deviate from traditional rational-choice predictions under different levels of social interactions. The emergence of prosocial decision making, defined as any action that is addressed to benefit another individual even at the expense of personal benefits, has been reported as an important example of such social influence. Furthermore, brain evidence has shown the involvement of structures such the prefrontal cortex, anterior insula and midcingulate cortex during decision settings in which a decision maker interacts with others under physical pain or distress or while being observed by others. Using a slightly modified version of the dictator game, we tested the hypothesis that the inclusion of another person into the decision setting increases prosocial decisions in young adults and that this increase is higher when the other person is associated with others in need. At the brain level, we hypothesized that the increase in prosocial decisions correlates with frontal theta activity as a marker of empathy saliency. The results showed that the inclusion of another person into the decision setting increased prosocial behavior only when this presence was associated with someone in need and that this was associated with an increase in frontocentral theta-oscillatory activity. These results suggest that the presence of someone in need enhances both empathy concerns and norm compliance, raising the participants’ prosocial decision making.
... Finally, the source current strength (3 x5,005 vertices of the cortex surface) is obtained by multiplying the recorded EEG signal at each electrode on the time series by the wMNE inverse operator. Importantly, this linear transformation allows the time-frequency analysis to be calculated directly on the source space without changing the spectral characteristics of the underlying source (Ambrosini & Vallesi, 2016;Billeke et al., 2013). Z-score transformations were conducted to normalize the theta source results after averaging all the trials for each condition, using the 500 to −200 ms pre-feedback baseline as the reference interval. ...



Pursuing dating relationships is important for many people's well-being, because it helps them fulfill the need for stable social relationships. However, the neural underpinnings of decision-making processes during the pursuit of dating interactions are unclear. In the present study, we used a novel online speed dating paradigm where participants (undergraduate students, N = 25, aged 18-25 years, 52% female) received direct information about acceptance or rejection of their various speed dates. We recorded EEG measurements during speed dating feedback anticipation and feedback processing stages to examine the stimulus preceding negativity (SPN) and feedback-related brain activity (Reward Positivity, RewP, and theta oscillatory power). The results indicated that the SPN was larger when participants anticipated interest versus disinterest from their speed dates. A larger RewP was observed when participants received interest from their speed dates. Theta power was increased when participants received rejection from their speed dates. This theta response could be source-localized to brain areas that overlap with the physical pain matrix (anterior cingulate cortex, dorsolateral prefrontal cortex, and the supplementary motor area). This study demonstrates that decision-making processes-as evident in a speed date experiment-are characterized by distinct neurophys-iological responses during anticipating an evaluation and processing thereof. Our results corroborate the involvement of the SPN in reward anticipation, RewP in reward processing and mid-frontal theta power in processing of negative social-evaluative feedback. These findings contribute to a better understanding of the neurocognitive mechanisms implicated in decision-making processes when pursuing dating relationships.
... They bias input selection, influence sensory processing, and support information transfer between cortical areas (18). Importantly, specific frequencies of neural oscillations have been implicated in decision making (e.g., b-band, [12][13][14][15][16][17][18][19][20][21][22][23][24]19), attention and stimulus selection (e.g., a-band, [8][9][10][11][12]20), and reflect arousal (e.g., aand b-band; 21 and 22). With respect to risk perception, an EEG study found a reduction of a-band power before feedback in social highrisk decisions (23). ...
... Importantly, specific frequencies of neural oscillations have been implicated in decision making (e.g., b-band, [12][13][14][15][16][17][18][19][20][21][22][23][24]19), attention and stimulus selection (e.g., a-band, [8][9][10][11][12]20), and reflect arousal (e.g., aand b-band; 21 and 22). With respect to risk perception, an EEG study found a reduction of a-band power before feedback in social highrisk decisions (23). However, little is known about the spectral fingerprints of the cortical correlates of risk-sensitive SCRs. ...
... In turn, reduced a-band power is related to the processing of relevant information: reduced a-power allows to sequentially process bits of information and prioritize the resulting objects according to salience, which can then capture attention based on subjective relevance (20). A study on social decisions found that a-band power was reduced before feedback in high-risk situations, suggesting a sensitivity to risk perception (23). In our experiment, before an electric shock, we observed reduced power in the frequency range between 3 and 17 Hz, which comprises the h (4-7 Hz), a (8-12 Hz), and b (13)(14)(15)(16)(17)(18)(19)(20)(21)(22)(23)(24)(25)(26)(27)(28)(29)(30) ranges, but appears to be maximal for the a-band (Fig. 4A). ...



Understanding the neural correlates of risk-sensitive skin conductance responses can provide insights into their connection to emotional and cognitive processes. To provide insights into this connection, we studied the cortical correlates of risk-sensitive skin conductance peaks using electroencephalography. Fluctuations in skin conductance responses were elicited while participants played a threat-of-shock-card-game. Precise temporal information about skin conductance peaks were obtained by applying continuous decomposition analysis on raw electrodermal signals. Shortly preceding skin conductance peaks, we observed a decrease in oscillatory power in the frequency range between 3 and 17 Hz in occipitotemporal cortical areas. Atlas-based analysis indicated the left lingual gyrus as the source of the power decrease. The oscillatory power averaged across 3 to 17 Hz showed a significant negative relationship with the skin conductance peak amplitude. Our findings indicate a possible interaction between attention and threat perception.
... Gehring et al. (2012) extracted theta oscillations using the Morlet wavelet transform and found frontally focused theta (4-7 Hz) activity for monetary losses compared with monetary gains when the medial frontal negativity (MFN) was evoked. Billeke et al. (2013) found that theta activity, which reflects the activity of dACC, was modulated by the individual strategy of the study participant. Telpaz et al. (2015) found a correlation between the theta band power and participants' rank-ordered preferences without requiring any hemispheric asymmetry. ...
Recent studies have suggested that event-related brain potential (ERP) can represent consumer preference, and there is consensus that the N200 is the best indicator of consumer preference. Measurement of reference-dependent consumer preference, in turn, requires a reference point, but it remains largely unknown how reference points modulate the preference-related N200. We designed an experiment to investigate how reference points affect the N200 based on classical paradigms. In the single-reference condition, one product was displayed in each trial; in the conjoined-reference condition, a pair of products was displayed simultaneously. Our results showed that in the single-reference condition, low-preference products elicited more negative N200 than high-preference products, replicating previous results, but the N200 could not distinguish between low‐ and high-preference products when viewing two options of similar subjective value in the conjoined-reference condition. These findings suggest that reference points modulate the representation of the N200 on consumer preference. When only viewing one product, participants make a value judgment based on their expectations. However, when viewing two products simultaneously, both their expectation and the alternative product can serve as reference points, and whether the N200 can represent consumer preference depends on which reference point is dominant. In future research, reference points must be controlled when the N200 is used to explore value-related decision-making.