Table 1 - uploaded by Spencer Craig Brown
Content may be subject to copyright.

Source publication









The legume genus Aeschynomene is notable in the ability of certain semiaquatic species to develop nitrogen‐fixing stem nodules. These species are distributed in two clades. In the first clade, all the species are characterized by the use of a unique Nod‐independent symbiotic process. In the second clade, the species use a Nod‐dependent symbiotic pr...
Contexts in source publication
Context 1
... the accessions of Aeschynomene used in this study, their geo- graphic origin and their sources are listed in Table 1 and Sup- porting Information Table S1. Seeds were scarified with sulfuric acid for germination and plants were grown in pots filled with atapulgite under glasshouse conditions (temperature, 26-36°C; relative humidity, 70-80%; insect-proof screens) as detailed in Arrighi et al. (2012). ...
Context 2
... investigate the characteristics of the whole group, we increased the taxonomic sampling performed by Chaintreuil et al. (2013) and used for comparison the diploid A. evenia as a representative of the Nod-independent group that was previously analyzed in Arrighi et al. (2014) (Fig. 1). A total of 38 accessions were sampled, procured from either different seed banks or direct field collections ( Table 1). All come from the African continent, with the exception of A. aspera which is exclusively Asian. ...
Context 3
... observed in our collection of accessions two flower morphotypes with signifi- cantly different sizes, being either c.16-20 mm or c. 24 mm long. To distinguish them, accessions displaying the largest flowers were qualified as 'grandiflora', as previously proposed by Peter (1928) (Table 1). ...
Context 4
... contents were estimated by flow cytometry for most accessions in order to provide a first view of the characteristics of the genomes encountered in the African group (Table 1). When comparing the species, genome size was found to present an extended range, varying from 1.92 pg/2C for A. cristata to 5.28 pg/2C for A. afraspera and up to 9.20 pg/2C for some A. schimperi accessions. ...
Context 5
... genome sizes may be indicative of different genome structures underlined by ploidy changes and generally larger genomes for the whole African group. It is also worth noting that for A. schimperi, the 'grandiflora' morphotype displays a 3.9 pg/2C genome size while other accessions are endowed with a 4.8, 8.5 or 9.2 pg/2C genome (Table 1). ...
Context 6
... link the genome sizes to ploidy levels, some accessions for each species were selected for chromosome counting (Table 1; Fig. S1). Five chromosome numbers were obtained: 2n = 28 for A. schimperi 'grandiflora' and one A. schimperi accession; 2n = 38 for A. aspera, A. fluitans, A. nilotica and A. cristata;2 n = 40 for A. crassicaulis, A. elaphroxylon, A. pfundii and A. uniflora;2 n = 56 for other A. schimperi accessions; and 2n = 76 for A. afraspera. ...
Context 7
... test these hypotheses regarding the evolution of chromosome numbers in the African Aeschynomene, the genetic relationships between the different species was investigated. For this, phyloge- netic reconstruction of the whole group was undertaken using the nuclear ribosomal ITS region and the chloroplast matK gene (Tables S1, S2). The maximum likelihood analysis of these two sequences produced very similar topologies (Fig. 2a,b) resolving the accessions in five well-supported clades (BS > 74% for ITS and BS > 98% for matK) with coherent chromosome numbers. ...
Context 8
... markers producing the same amplification profile are listed below the picture and the one used for illustration is underlined. Numbers following the species names refer to the accession code shown in Table 1. ...
Context 9
... further substantiate the hypothesis of a hybrid origin, we took advantage of a transcriptome database available for A. afraspera (Czernic et al., 2015) to develop a set of 36 AaSSR markers (Table S1) and to perform cross-amplification tests on the species related to A. afraspera, using one accession per species. Fifteen AaSSR markers amplified in the five accessions tested, whereas 21 AaSSR markers displayed restricted amplification profiles. ...
Context 10
... in silico analysis revealed three to four distinct sequences for many genes for A. afraspera when only one sequence was encountered for A. evenia that is known to be diploid. Six genes were selected for detailed comparisons: ascor- bate peroxidase 2 (APX2), asparagine synthetase 2 (AS2), eukary- otic translation initiation factor a (eiF1a), plasma membrane intrinsic protein 2;7 (PIP2;7), sucrose synthase (SUS) and tono- plast intrinsic protein 1;1 (TIP1;1) genes (Table S1). Phyloge- netic analyses of the coding sequences of the six single-copy nuclear genes by the maximum likelihood method always resulted in a topology that clustered the distinct sequences into two highly divergent clades (BS of 87-100%). ...
Context 11
... the reconstruction of the polyploidization history of A. afraspera in line with the other members of the African clade, we selected two genes, eif1a and SUS, being either intronless or presenting a long exon, and sequenced them in selected acces- sions representing the different species of the group. To capture all possible copies of these genes, several primers were developed based on the transcriptome sequences of A. afraspera (Table S1). In most cases, we isolated a pair of putative homeologs, and in several other instances three or four putative homeologs. ...
Context 12
... afraspera is hypothesized to result from the hybridization of A. cristata and another genome donor related to A. fluitans and A. nilotica, based on phylogenetic analyses. In clade 1, A. cristata is notable in that it displays a relatively small genome (1.92 pg/2C) that corresponds to only 46% of the genome size of three other 4x species, A. aspera, A. fluitans and A. nilotica (Table 1). Thus, A. cristata shows a drastic and specific genome downsizing. ...
Context 13
... schimperi (8x) is also supposed to derive from hybridization between A. schimperi (4x) and A. schimperi 'grandi- flora' (4x). As these putative progenitors present a 20% difference in their genome size (4.82 and 3.93 pg/2C, respectively) (Table 1), a similar cytogenetic approach was performed. Metaphase chromosomes of A. schimperi (4x) appeared relatively longer than those of A. schimperi 'grandiflora', but most strikingly they presented an atypical plump morphotype and well- developed telomeres (Fig. 4) ...
Citations
... The photosynthetic ability of Aeschynomene-nodulating bradyrhizobia was believed to be similar to that in cyanobacteria and a number of other bacterial groups [5], while all of the photosynthesis species in Bradyrhizobium have been classified into the photosynthetic supergroup [5,6], corresponding to the dominant symbionts (B. oligotrophicum, B. denitrificans, and B. aeschynomenes) of Aeschynomene species [5,7,8]. Previously, a complex evolutionary history of photosynthesis has been found in Bradyrhizobium strains [9], and Bradyrhizobium sp. ...

Aeschynomene indica rhizobia (AIRs) are special classes of bacteria capable of nodulating without nodulation factors and have photosynthetic capacity. With an aim to characterize the structural variations in Bradyrhizobium genomes during its evolution, the genomes of AIRs and the reference Bradyrhizobium strains were compared utilizing molecular biology, bioinformatics, and biochemistry techniques. The presence of symbiotic nitrogen fixation (nif) genes and photosynthetic genes, as well as components of the T3SS (Type III secretion system) and T3CP (Type III chaperone) in the genome of AIRs, was also assessed. Additionally, the origin, evolutionary history, and genes associated with the NF-independent nodulation pattern in AIRs were explored. The results indicate that horizontal gene transfer events have occurred in AIRs, and three distinct origins of AIRs were estimated: early differentiated AIRs, non-symbiotic Bradyrhizobium, and non-AIRs. In contrast to the significant genetic transformations observed in the second and third groups, the first group of AIRs displays a rich evolutionary history, exhibits high species diversity, and primarily relies on vertical transmission of nitrogen fixation and photosynthetic genes. Overall, the findings provide a fundamental theoretical foundation for gaining a deeper understanding of the phylogeny and genealogy of AIRs.
... www.nature.com/scientificreports/ correspond to the 8 × cytotype 9 . In non-waterlogged conditions, numerous pink nodules could be observed on the main root, while under flooded conditions (9 of the 12 sampled sites) green nodules were present in the lower part of the stem (Fig. 3b). ...
... uniflora (Fig. 1b) 26 . ITS sequencing for the samples JFA51 and JFA71 (examples of plants with prominent and small flowers, respectively) revealed differences in their sequences (5 single nucleotide polymorphisms (SNPs)) that seem to discriminate large flower A. uniflora accessions (including JFA51) from small flower-bearing ones presently and previously characterized (JFA71, LSTM61, LSTM137) ( Supplementary Fig. 2) 9 . Variations in the matK sequences were also observed but these were not correlated with the flower type ( Supplementary Fig. 2). ...
... Variations in the matK sequences were also observed but these were not correlated with the flower type ( Supplementary Fig. 2). Moreover, flow cytometry measurements were relatively homogeneous (, Supplementary Table 2) 9 . In both morphotypes, numerous green nodules with a spherical shape and a narrow neck, running on the stem were visible (Fig. 3c). ...



Legumes have the ability to establish a nitrogen-fixing symbiosis with soil rhizobia that they house in specific organs, the nodules. In most rhizobium-legume interactions, nodulation occurs on the root. However, certain tropical legumes growing in wetlands possess a unique trait: the capacity to form rhizobia-harbouring nodules on the stem. Despite the originality of the stem nodulation process, its occurrence and diversity in waterlogging-tolerant legumes remains underexplored, impeding a comprehensive analysis of its genetics and biology. Here, we aimed at filling this gap by surveying stem nodulation in legume species-rich wetlands of Madagascar. Stem nodulation was readily observed in eight hydrophytic species of the legume genera, Aeschynomene and Sesbania, for which significant variations in stem nodule density and morphology was documented. Among these species, A. evenia, which is used as genetic model to study the rhizobial symbiosis, was found to be frequently stem-nodulated. Two other Aeschynomene species, A. cristata and A. uniflora, were evidenced to display a profuse stem-nodulation as occurs in S. rostrata. These findings extend our knowledge on legumes species that are endowed with stem nodulation and further indicate that A. evenia, A. cristata, A. uniflora and S. rostrata are of special interest for the study of stem nodulation. As such, these legume species represent opportunities to investigate different modalities of the nitrogen-fixing symbiosis and this knowledge could provide cues for the engineering of nitrogen-fixation in non-legume crops.
... In Bradyrhizobium, photosynthesis is thought to be limited to the so-called 'Photosynthetic supergroup' [17,23] whose members (e.g. B. oligotrophicum and B. denitrificans) are predominantly symbionts of species in the Aeschynomene genus of aquatic legumes [23][24][25]. ...




Bradyrhizobium comprises a diverse group of bacteria with various lifestyles. Although best known for their nodule-based nitrogen-fixation in symbiosis with legumes, a select group of bradyrhizobia are also capable of photosynthesis. This ability seems to be rare among rhizobia, and its origin and evolution in these bacteria remain a subject of substantial debate. Therefore, our aim here was to investigate the distribution and evolution of photosynthesis in Bradyrhizobium using comparative genomics and representative genomes from closely related taxa in the families Nitrobacteraceae, Methylobacteriaceae, Boseaceae and Paracoccaceae . We identified photosynthesis gene clusters (PGCs) in 25 genomes belonging to three different Bradyrhizobium lineages, notably the so-called Photosynthetic, B. japonicum and B. elkanii supergroups. Also, two different PGC architectures were observed. One of these, PGC1, was present in genomes from the Photosynthetic supergroup and in three genomes from a species in the B. japonicum supergroup. The second cluster, PGC2, was also present in some strains from the B. japonicum supergroup, as well as in those from the B. elkanii supergroup. PGC2 was largely syntenic to the cluster found in Rhodopseudomonas palustris and Tardiphaga . Bayesian ancestral state reconstruction unambiguously showed that the ancestor of Bradyrhizobium lacked a PGC and that it was acquired horizontally by various lineages. Maximum-likelihood phylogenetic analyses of individual photosynthesis genes also suggested multiple acquisitions through horizontal gene transfer, followed by vertical inheritance and gene losses within the different lineages. Overall, our findings add to the existing body of knowledge on Bradyrhizobium ’s evolution and provide a meaningful basis from which to explore how these PGCs and the photosynthesis itself impact the physiology and ecology of these bacteria.
... Polyploidy is a common phenomenon in legumes (Doyle 2012;Cannon et al. 2015;Doyle and Sherman-Broyles 2017). Within dalbergioids, multiple species of Arachis L. (Pterocarpus clade; Bertioli et al. 2016), Stylosanthes (Pterocarpus clade; Marques et al. 2018), and Aeschynomene (Dalbergia clade; Arrighi et al. 2014;Chaintreuil et al. 2013Chaintreuil et al. , 2016Brottier et al. 2018) are polyploids. However, the precise estimation of the polyploid frequency in a particular group depends on the clear definition of the base chromosome number (x) (Barker et al. 2016;Doyle and Sherman-Broyles 2017). ...
... Phylogenetic analysis -To build the phylogenetic tree for this study, we retrieved 154 chloroplast matK gene and flanking trnK intron sequences representing 152 species from 44 genera of dalbergioids (the only genus from dalbergioid legumes not represented here is Maraniona C.E. Hughes, G.P. Lewis, Daza & Reynel). The species sequences were mainly published by Lavin et al. (2001), Chaintreuil et al. (2016), andBrottier et al. (2018), and a few sequences were selected directly from GenBank (Online Resource 1). We chose the species aiming to optimize that all species with published chromosome numbers and available sequences were represented in the phylogenetic tree; however, we did not include all available sequences for all species (see Online Resource 1). ...
... Phylogenetic relationships among dalbergioid legumes The tree topologies obtained in this study, from ML and BI, were nearly identical and similar to those of previous works (Lavin et al. 2001;Wojciechowski et al. 2004;Cardoso et al. 2013;Chaintreuil et al. 2016;Brottier et al. 2018). Here we sampled 44 of the 45 dalbergioid genera and almost all dalbergioid genera were recovered as monophyletic, except Aeschynomene and Humularia which is nested inside A. subg. ...



The dalbergioids, one of the largest clades among legumes, present significant karyotype diversity. Previous studies suggested different base chromosome numbers, hampering the understanding of karyotype evolution in dalbergioid legumes. In this study, we integrated chromosome numbers from the literature with phylogenetic data and new cytogenetic data for six species aiming to clarify the paths of karyotype evolution. We could confirm the base chromosome number of dalbergioid legumes as x = 10, which was constant for the three dalbergioid clades, namely Adesmia, Dalbergia, and Pterocarpus. However, we found alternative base chromosome numbers for some genera, which illustrate the two main mechanisms of chromosome changes found in dalbergioids: polyploidy and dysploidy. Considering the first diverging lineages in the phylogeny, polyploidy and dysploidy could be detected in seven and three nodes, respectively, with two out of the three dysploidy events associated with polyploidy. Nevertheless, considering the chromosome changes within species, the descendent dysploidy was the most common event in dalbergioid legumes. Such a high frequency of dysploidy highlights the importance of studying multiple populations from each species. In addition, the chromosome banding revealed DAPI+ bands, which is not commonly found in plants, as a possible chromosome marker for Stylosanthes. The chromosome banding analysis has taxonomic potential and could be explored in future studies to better understand the complex taxonomic groups inside dalbergioid legumes.
... The genus has also been shown to be paraphyletic, a number of related genera being nested within, but altogether they form a distinct clade in the tribe Dalbergieae [4,[23][24][25][26]. Within this broad clade, two groups of semi-aquatic Aeschynomene have been well-studied from a genetic and genomic standpoint: the A. evenia group, which contains all the Nod-independent species (most of them being 2x), and the A. afraspera group (all species being Nod-dependent) that appears to have a 4x origin [27][28][29]. For comparative analyses, the use of Nod-dependent species with a diploid structure would be more appropriate, but such Aeschynomene species are poorly documented. ...
... This strategy allowed checking the species identity and obtaining complementary data on the same plant material. DNA was isolated for 40 out of the 41 species (compared to the 27 species used in [4]) included in this group in taxonomic and genetic studies (Additional file 1: Table S1) [4,21,[27][28][29]. In addition, to determine the phylogenetic relationship of this subgenus with Aeschynomene subgenera Bakerophyton and Rueppellia, unclassified Aeschynomene species, as well as with the allied genera Bryaspis, Cyclocarpa, Geissaspis, Humularia, Kotschya, Smithia and Soemmeringia, we also sampled all these 10 taxa (compared to the 5 taxa present in [4]) [23,30]. ...
... Previous studies had demonstrated that the A. evenia clade is mostly diploid (2n = 2x = 20) even if some species such as A. indica (2n = 4x = 40, 2n = 6x = 60) appear to be of recent allopolyploid origin [27,29]. Conversely, all the species of the A. afraspera group were found to be polyploid (2n = 4x = 28,38,40, 2n = 8x = 56,76) and to have a common AB genome structure but the origin of the polyploidy event remained undetermined [28]. To assess the ploidy levels in Aeschynomene species and related genera, chromosome numbers and nuclear DNA content were determined (appended to labels in Fig. 2 a, Additional file 1: Table S1, Additional file 6: Figure S3 and Additional file 7: Figure S4). ...

Background
Among semi-aquatic species of the legume genus Aeschynomene, some have the property of being nodulated by photosynthetic Bradyrhizobium lacking the nodABC genes necessary for the synthesis of Nod factors. Knowledge of the specificities underlying this Nod-independent symbiosis has been gained from the model legume Aeschynomene evenia but our understanding remains limited due to the lack of comparative genetics with related taxa using a Nod factor-dependent process. To fill this gap, we combined different approaches to perform a thorough comparative analysis in the genus Aeschynomene.
Results
This study significantly broadened previous taxon sampling, including in allied genera, in order to construct a comprehensive phylogeny. In the phylogenetic tree, five main lineages were delineated, including a novel lineage, the Nod-independent clade and another one containing a polytomy that comprised several Aeschynomene groups and all the allied genera. This phylogeny was matched with data on chromosome number, genome size and low-copy nuclear gene sequences to reveal the diploid species and a polytomy containing mostly polyploid taxa. For these taxa, a single allopolyploid origin was inferred and the putative parental lineages were identified. Finally, nodulation tests with different Bradyrhizobium strains revealed new nodulation behaviours and the diploid species outside of the Nod-independent clade were compared for their experimental tractability and genetic diversity.
Conclusions
The extended knowledge of the genetics and biology of the different lineages sheds new light of the evolutionary history of the genus Aeschynomene and they provide a solid framework to exploit efficiently the diversity encountered in Aeschynomene legumes. Notably, our backbone tree contains all the species that are diploid and it clarifies the genetic relationships between the Nod-independent clade and the Nod-dependent lineages. This study enabled the identification of A. americana and A. patula as the most suitable species to undertake a comparative genetic study of the Nod-independent and Nod-dependent symbioses.
Electronic supplementary material
The online version of this article (10.1186/s12870-018-1567-z) contains supplementary material, which is available to authorized users.
... The genus has also been shown to be paraphyletic, a number of related genera being nested within, but altogether they form a distinct clade in the tribe Dalbergieae [4,[23][24][25][26]. Within this broad clade, two groups of semi-aquatic Aeschynomene have been well-studied from a genetic and genomic standpoint: the A. evenia group, which contains all the Nod-independent species (most of them being 2x), and the A. afraspera group (all species being Nod-dependent) that appears to have a 4x origin [27][28][29]. For comparative analyses, the use of Nod-dependent species with a diploid structure would be more appropriate, but such Aeschynomene species are poorly documented. ...
... This strategy allowed checking the species identity and obtaining complementary data on the same plant material. DNA was isolated for 40 out of the 41 species (compared to the 27 species used in [4]) included in this group in taxonomic and genetic studies (Additional file 1: Table S1) [4,21,[27][28][29]. In addition, to determine the phylogenetic relationship of this subgenus with Aeschynomene subgenera Bakerophyton and Rueppellia, unclassified Aeschynomene species, as well as with the allied genera Bryaspis, Cyclocarpa, Geissaspis, Humularia, Kotschya, Smithia and Soemmeringia, we also sampled all these 10 taxa (compared to the 5 taxa present in [4]) [23,30]. ...
... Previous studies had demonstrated that the A. evenia clade is mostly diploid (2n = 2x = 20) even if some species such as A. indica (2n = 4x = 40, 2n = 6x = 60) appear to be of recent allopolyploid origin [27,29]. Conversely, all the species of the A. afraspera group were found to be polyploid (2n = 4x = 28,38,40, 2n = 8x = 56,76) and to have a common AB genome structure but the origin of the polyploidy event remained undetermined [28]. To assess the ploidy levels in Aeschynomene species and related genera, chromosome numbers and nuclear DNA content were determined (appended to labels in Fig. 2 a, Additional file 1: Table S1, Additional file 6: Figure S3 and Additional file 7: Figure S4). ...
Some Aeschynomene legume species have the property of being nodulated by photosynthetic Bradyrhizobium lacking the nodABC genes. Knowledge of this unique Nod (factor)-independent symbiosis has been gained from the model A. evenia but our understanding remains limited due to the lack of comparative genetics with related taxa using a Nod-dependent process.
To fill this gap, this study significantly broadened previous taxon sampling, including in allied genera, to construct a comprehensive phylogeny. This backbone tree was matched with data on chromosome number, genome size, low-copy nuclear genes and strengthened by nodulation tests and a comparison of the diploid species.
The phylogeny delineated five main lineages that all contained diploid species while polyploid groups were clustered in a polytomy and were found to originate from a single paleo-allopolyploid event. In addition, new nodulation behaviours were revealed and Nod-dependent diploid species were shown to be tractable.
The extended knowledge of the genetics and biology of the different lineages in the legume genus Aeschynomene provides a solid research framework. Notably, it enabled the identification of A. americana and A. patula as the most suitable species to undertake a comparative genetic study of the Nod-independent and Nod-dependent symbioses.
... section SI) using this A. evenia (CIAT22838) FEN1 sequence as basis, we obtained four different sequences of putative FEN1 genes for A. afraspera and one sequence for another A. evenia line, PI 225551. This difference in the number of FEN1 homologs between these two species is not surprising considering that A. evenia is diploid whereas A. afraspera is octoploid 16 . Phylogenetic analysis showed that all the gene products clustered in a clade containing L. japonicus FEN1 (Fig. 5A). ...
... The procedure for the identification of FEN1 and IPMS orthologs in A. evenia (CIAT22838/PI 225551) and A. afraspera ((LSTM #1) is described in detail in the Supplementary information section. Using the obtained sequences a phylogenetic analysis was performed as described in 16 and data are presented as rooted trees using the Cucumis IPMS as outgroup. GenBank/EMBL and Gene_ID numbers for sequences obtained and used for phylogenetic analysis can be found in Table S4 of the Supplementary information section. ...



In the most studied rhizobium-legume interactions, the host plant supplies the symbiont with homocitrate, an essential co-factor of the nitrogenase enzyme complex, via the expression of a nodule-specific homocitrate synthase FEN1. Photosynthetic bradyrhizobia interacting with Nod factor (NF) dependent and NF-independent Aeschynomene legumes are able to synthesize homocitrate themselves as they contain a nifV gene encoding a homocitrate synthase. Here, we show that in the model strain ORS285, nifV is required for free-living and symbiotic dinitrogen fixation with NF-independent Aeschynomene species. In contrast, in symbiosis with NF-dependent Aeschynomene species, the nifV requirement for efficient nitrogen fixation was found to be host plant dependent. Interestingly, orthologs of FEN1 were found in both NF-dependent and NF-independent Aeschynomene species. However, a high nodule specific induction of FEN1 expression was only observed in A. afraspera, a host plant in which nifV is not required for symbiotic dinitrogen fixation. These data indicate that efficient symbiotic nitrogen fixation in many of the tested Aeschynomene species requires rhizobial homocitrate synthesis. Considering that more than 10% of the fully sequenced rhizobium strains do contain a nifV gene, the Aeschynomene/photosynthetic Bradyrhizobium interaction is likely not the only rhizobium/legume symbiosis where rhizobial nifV expression is required.
... Onge et al., 2012) and through phylogenetic analysis of orthologous transcripts in the conifer Sequoia sempervirens (Scott et al., 2016). Unfortunately, extensive genomic rearrangements after WGD, as well as more recent WGDs, can obscure the nature of an ancient WGD to a point where multiple lines of evidence either cannot resolve an auto-or allopolyploid origin or produce conflicting results (e.g., Chaintreuil et al., 2016). ...
Polyploidy (whole-genome duplication, WGD) is an integral feature of eukaryotic evolution with two main forms typically recognized, autopolyploidy and allopolyploidy. In plants, a growing body of research contradicts historical assumptions that autopolyploidy is both infrequent and inconsequential in comparison to allopolyploidy. However, the legacy of these assumptions still persists through a lack of research on central facets of autopolyploid evolution. This review highlights recent research that has significantly increased scientific understanding of autopolyploidy. Key advances include: 1) unreduced female gametes contribute disproportionally to polyploidization through the formation of triploids, 2) niche divergence in autopolyploids can occur immediately or gradually after WGD through a diverse set of mechanisms, but broad niche overlap is also common between diploids and autopolyploids, and 3) the degree of genomic and transcriptomic changes following WGD is lower in autopolyploids than allopolyploids, but is highly variable both within and between species in both types of polyploids. We discuss the implications of these and other recent findings, present promising systems for future research, and advocate for expanded research in diverse areas of autopolyploid evolution.
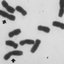
A cytogenetic analysis of sixteen taxa of the genus Aeschynomene Linnaeus, 1753, which includes species belonging to both subgenera Aeschynomene (Léonard, 1954) and Ochopodium (Vogel, 1838) J. Léonard, 1954, was performed. All studied species had the same chromosome number (2n = 20) but exhibited karyotype diversity originating in different combinations of metacentric, submetacentric and subtelocentric chromosomes, chromosome size and number of SAT chromosomes. The plasticity of the genomes included the observation in a taxon belonging to the subgenus Aeschynomene of an isolated spherical structure similar in appearance to the extra chromosomal circular DNA observed in other plant genera. By superimposing the karyotypes in a recent phylogenetic tree, a correspondence between morphology, phylogeny and cytogenetic characteristics of the taxa included in the subgenus Aeschynomene is observed. Unlike subgenus Aeschynomene, the species of Ochopodium exhibit notable karyotype heterogeneity. However the limited cytogenetic information recorded prevents us from supporting the proposal of their taxonomic separation and raise it to the genus category. It is shown that karyotype information is useful in the taxonomic delimitation of Aeschynomene and that the diversity in the diploid level preceded the hybridization/polyploidization demonstrated in the genus. The systematic implications of our results and their value can be extended to other Dalbergieae genera as knowledge about the chromosomal structure and its evolution increases.

En cas de carence azotée, les légumineuses sont capables de mettre en place une symbiose avec des bactéries du sol fixatrices d’azote appelées rhizobia. Cette symbiose a lieu dans un organe appelé nodosité où les bactéries sont endocytées et appelées bactéroïdes. Certains clades de légumineuses imposent un processus de différenciation à leurs bactéroïdes qui agrandissent considérablement et deviennent polyploïdes, menant à des morphotypes bactériens allongés ou sphériques. Au cours de cette thèse, j’ai étudié la différenciation des bactéroïdes de Bradyrhizobium spp. en association avec Aeschynomene spp.. Les bactéroïdes de ces plantes présentent des degrés de différenciation distincts qui dépendent de l’espèce hôte. Mes données suggèrent que les bactéroïdes les plus différenciés sont aussi les plus efficaces. J’ai cherché à savoir quels facteurs procaryotes pourraient être impliqués dans les adaptations des bactéroïdes au processus de différenciation et à leurs divers hôtes, le tout en lien avec cette différence d’efficacité symbiotique au travers d’approches globales sans a priori de type -omiques. Les conditions considérées sont des bactéroïdes de différents morphotypes et des cultures libres de référence. Les fonctions activées en conditions symbiotiques ont été identifiées, ainsi que les gènes spécifiques d’un hôte donné. Des analyses fonctionnelles des gènes d’intérêt ont également été menées. Les mutants bactériens n’ont toutefois pas présenté de phénotype symbiotique drastique, montrant ainsi l’existence de réseaux de gènes complexes menant à la résilience des génomes de rhizobia.